| |
Introduction
: the main bones |
| |
|
|
For a
complete description of the skeleton and muscles, specialized works such
as the Atlas d'Anatomie du Lapin, published by Barone et al. (1973) are
advised (Latin vocabulary, French and English). Also, articles such as
those written by Cantier et al. (1974), establish correspondence between
the names of the skeletal muscles adopted by different authors. |
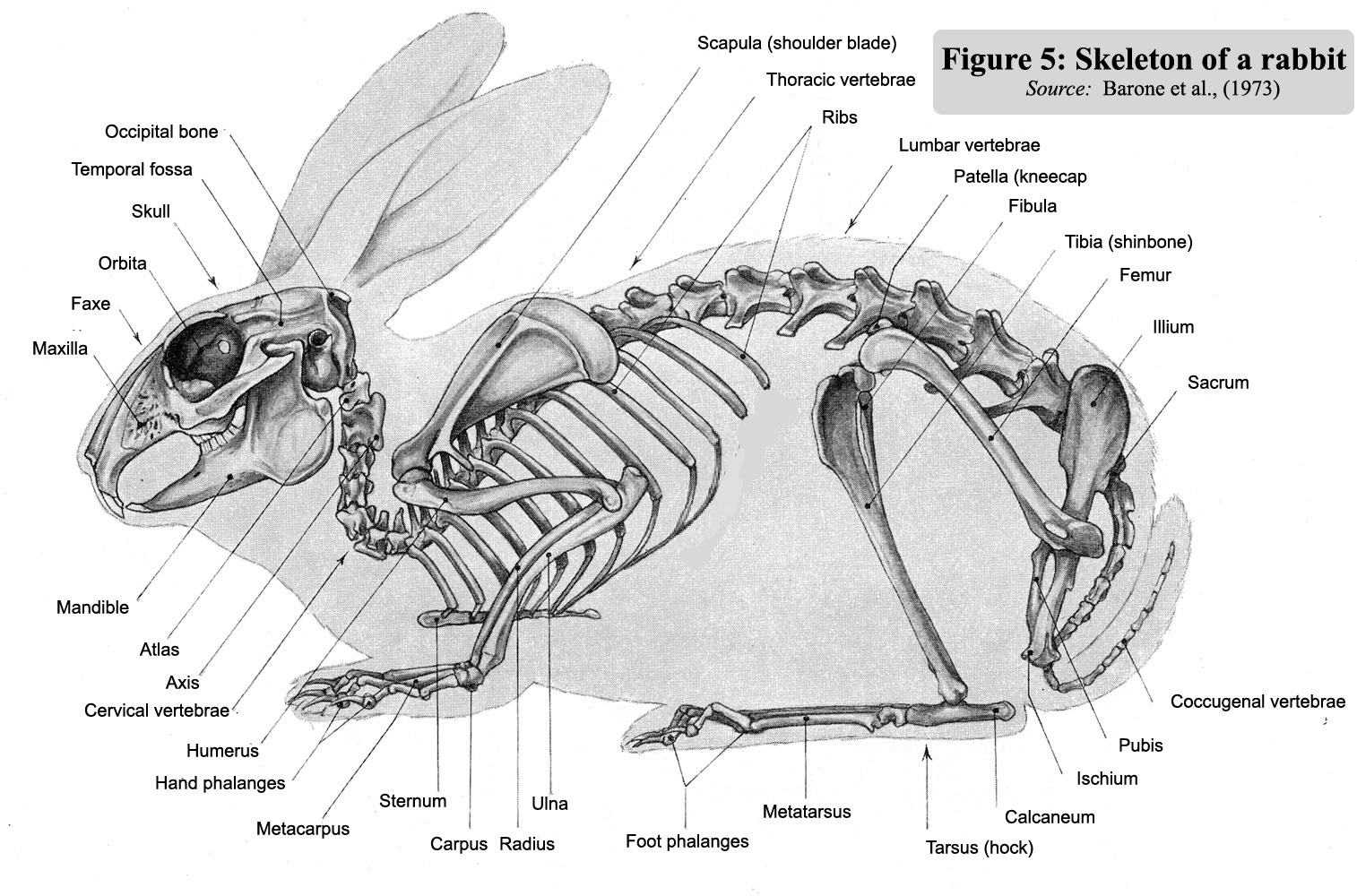
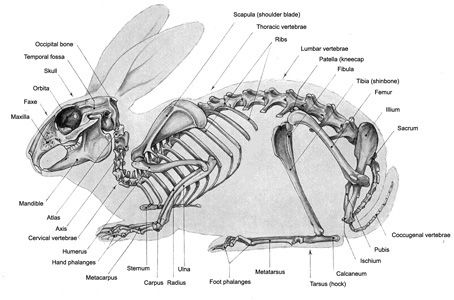 Figure 5 : Skeleton
of a rabbit (Soucre
: Barone et al., 1973)
Figure 5 : Skeleton
of a rabbit (Soucre
: Barone et al., 1973) |
| |
Skeleton
bones |
|
The
main bones appear in figure 5 opposite with names =>
Below example of a rabbit skeleton presentation (figure 5a)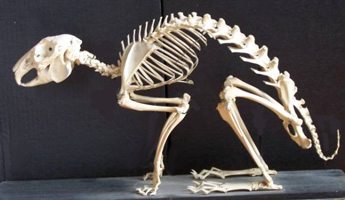
|
| |
Skull
and dentition |
|
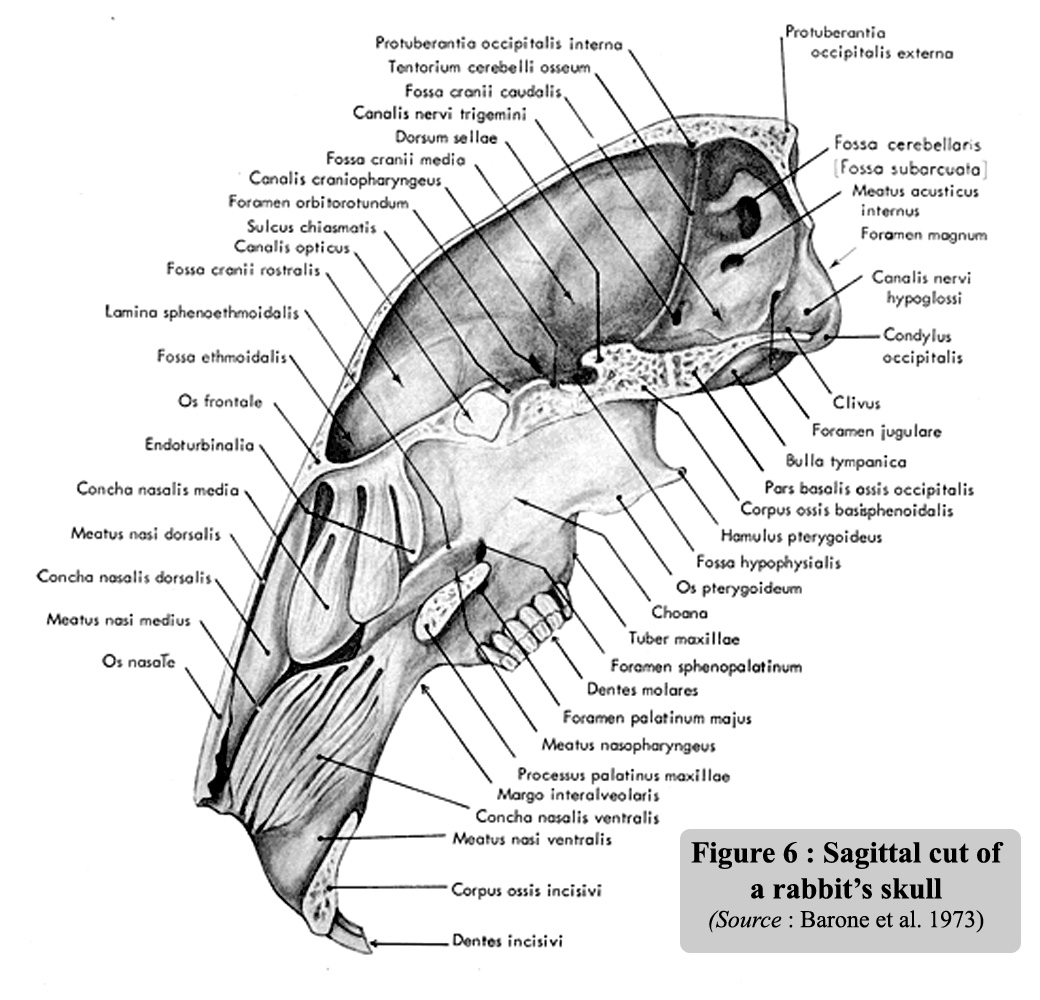
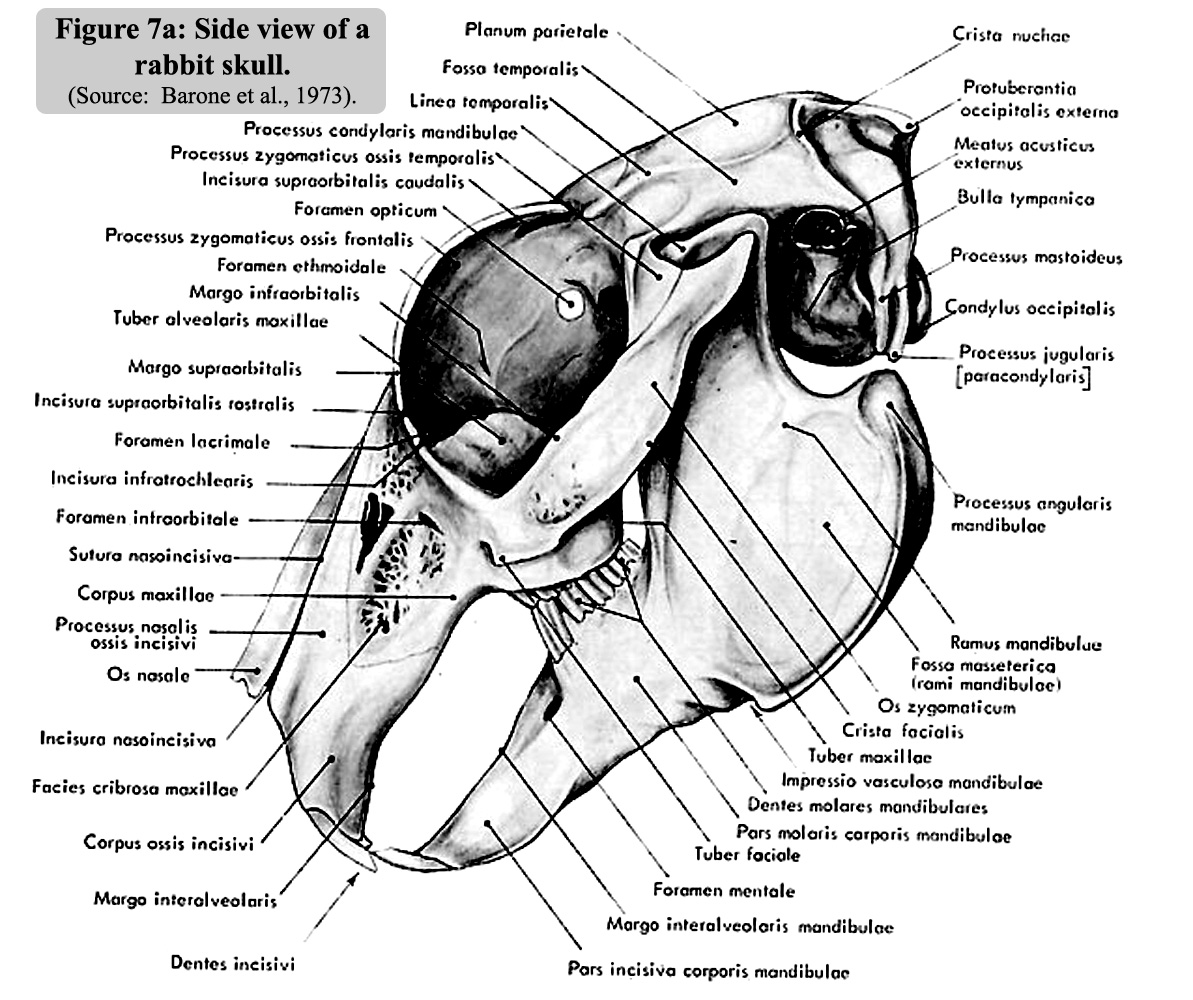
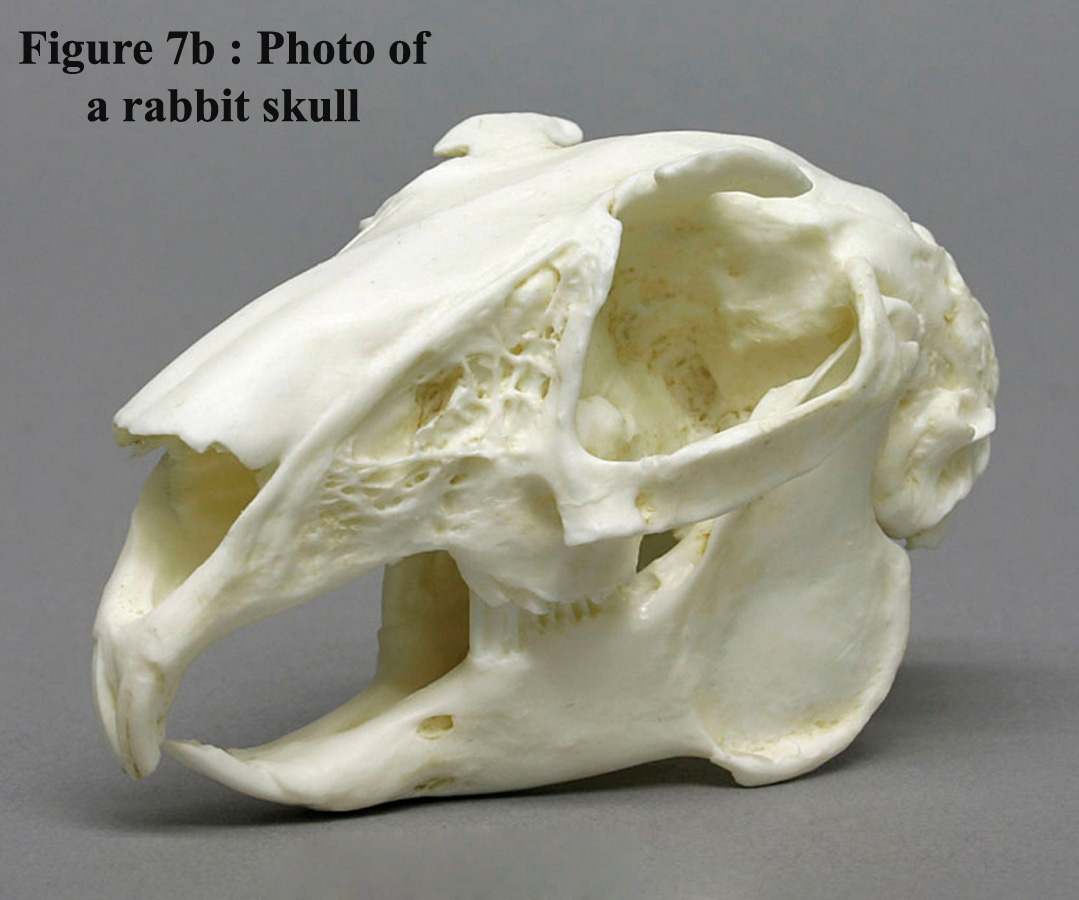
Figure
6 is the sagittal section of the skull. The size of the nasal sinuses,
which occupy one third of the volume of the inside of the skull, is worth
mentioning. A side view of the skull is illustrated in Figure 7a and a
photo of a rabbit skull is give on figure 7b |
| |
|
|
|
|
|
|
|
Figure
6: Sagittal cut of a rabbit's skull.
(
Source:
Barone et al., 1973).
|
Figure
7a: A side view of the skull.
(Source: Barone
et al., 1973).
|
Figure
7b : Photo of the skull of a rabbit
|
|
| |
|
|
|
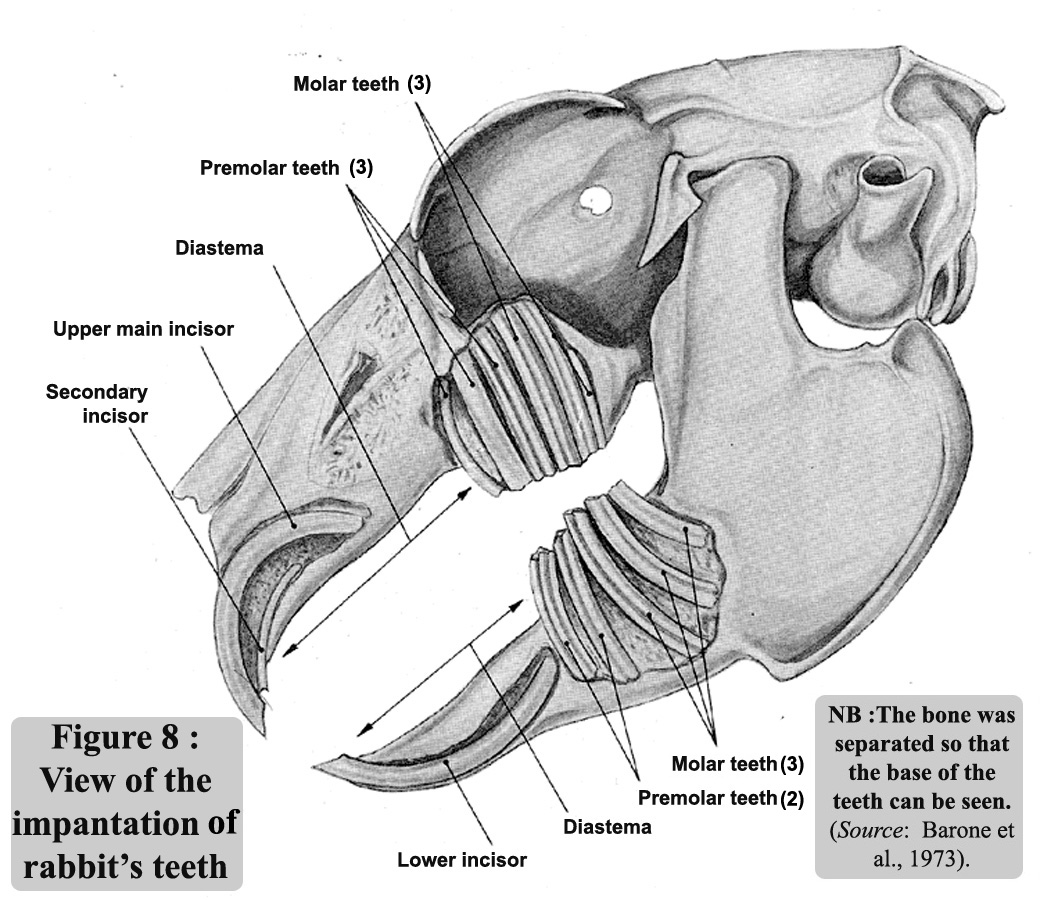
Figure 8 shows
the implantation of the teeth. Rabbits have two pairs of incisors
in the maxilla, and one pair in the mandible which has enabled
zoologists to clearly distinguish between rabbits (and lagomorphs
in general), and rodents that only have one pair in the maxilla
and one pair in the mandible. In the rabbit, the second pair of
incisors is situated behind the first and therefore, completely
hidden.
All incisors are covered with a layer of enamel which is thinner
at the back than the front, thus allowing the rabbit to sharpen
its teeth by filing the upper ones against the lower ones. The
anterior face has a longitudinal furrow.
There are no canines and a strong diastema separates the incisors
from the premolars (3 + 2 pairs) and molars (3 + 3 pairs).
In all, there are 2
pairs of incisors, 3 pairs of premolars, 3 pairs of molars in
the maxilla and 1 pair of incisors, 2 pairs of premolars and 3
pairs of molars in the mandible making a total of 28 teeth, of
which, 26 are only functional. As in the case of all lagomorphs,
the teeth are deeply embedded in the bones but have no roots (figure
8).
Their growth is continuous throughout the animal's life.
A measurement of the growth rate of the incisors gave a value
of 2 mm per week for the maxilla and 2.4 mm for the mandible.
That of the molars and premolars (also called "jugal"
teeth) is much slower and has been estimated at only 2 mm per
month (~ 4 times slower) (Shadle, 1936).
The kits are
born with milk teeth (incisors and premolars), which fall out
at around 18 days of age and are almost immediately replaced by
the permanent ones.
|
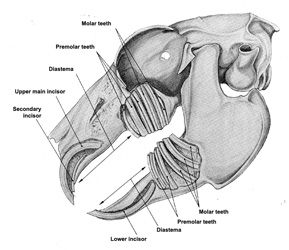 Figure 8
: View of the implantation of rabbit`s teeth. The bone was separated
so that the base of the teeth can be seen
Figure 8
: View of the implantation of rabbit`s teeth. The bone was separated
so that the base of the teeth can be seen. (Source:
Barone et al., 1973). |
|
| |
The
vertebrae and bones of the limbs |
| |
|
|
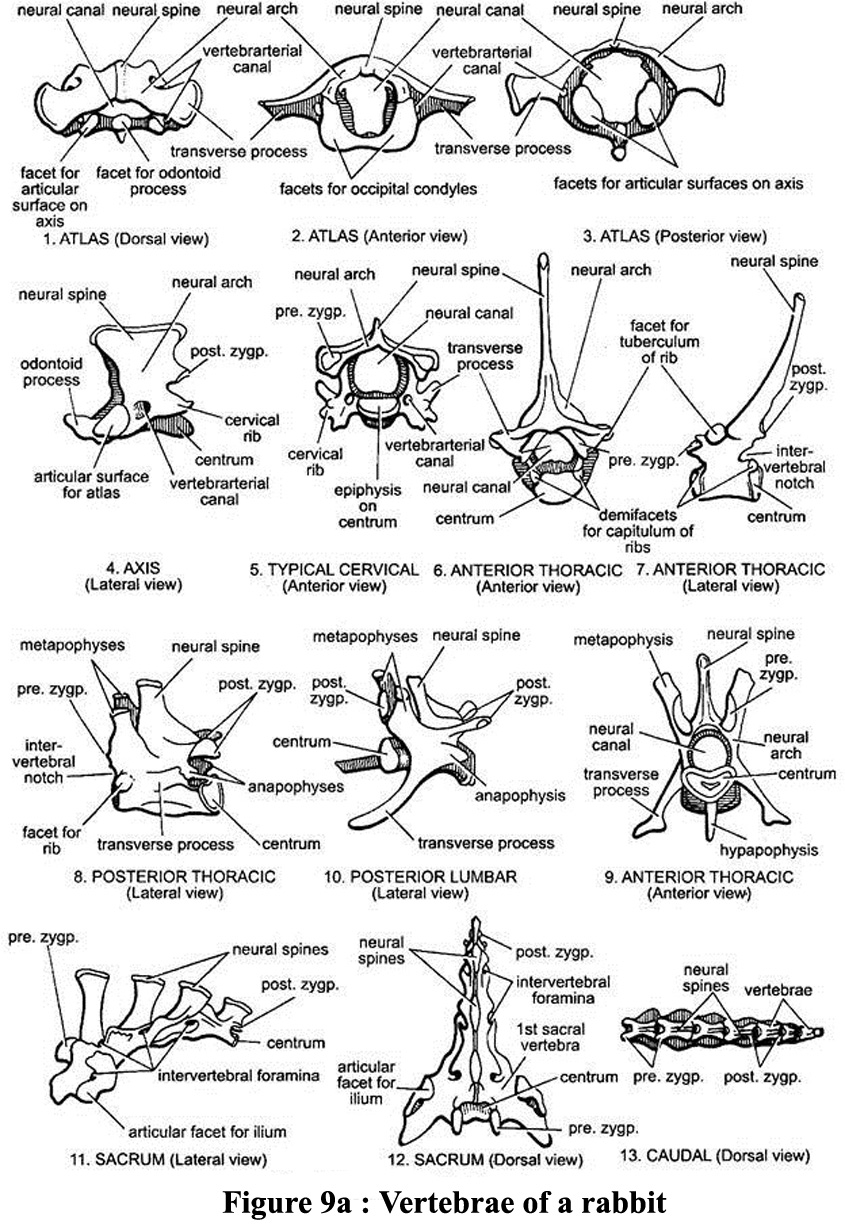
The skeleton has
7 cervical vertebrae, of which the first two are the atlas (carrying
the head) and the axis which plays a primordial role in the rotation
of the head. The 12 thoracic vertebrae are joined to 12 ribs.
Only the first 10 ribs are ventrally attached to the sternum to form
the thoracic cage. The other two ribs are called "floating ribs".
The 7 lumbar vertebrae are followed by the sacrum which
consists of 4 fused sacra vertebrae. The sacrum contains the pelvic
bones. Immediately after there are 15 coccygeal vertebrae, the
last 10 of which form the tail.. General organisdation of vertebrae
is descibed in figure 5. Detailed illustration of vertebral bone is
available on figure 9a.
|
| |
|
|
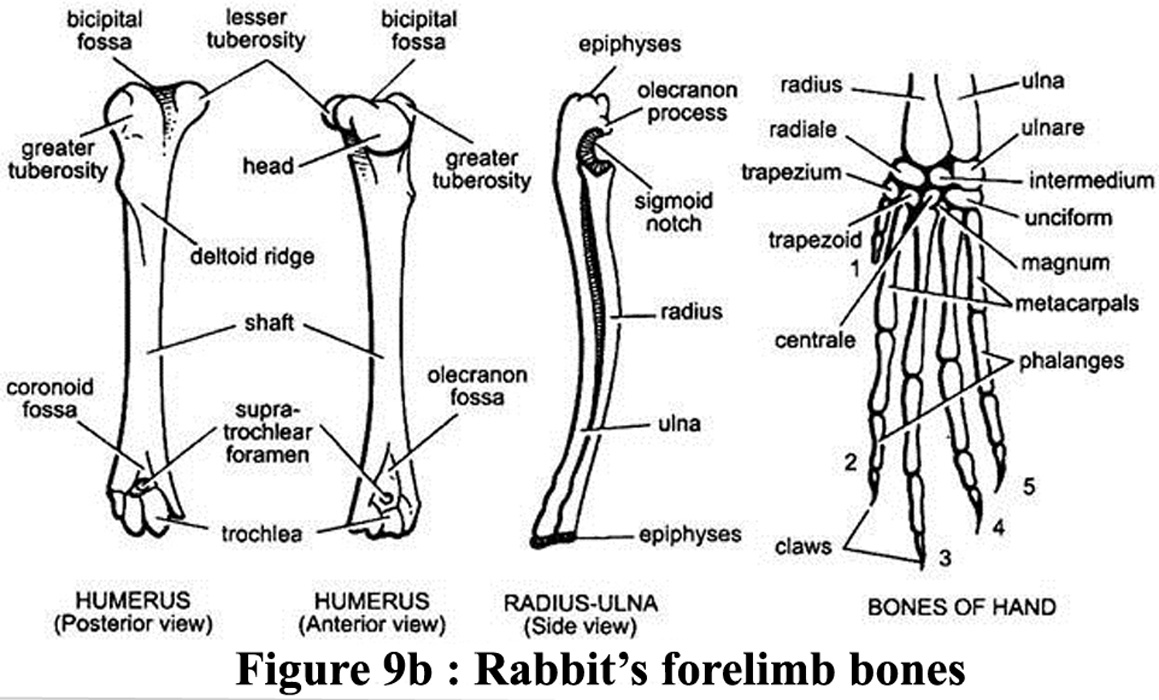
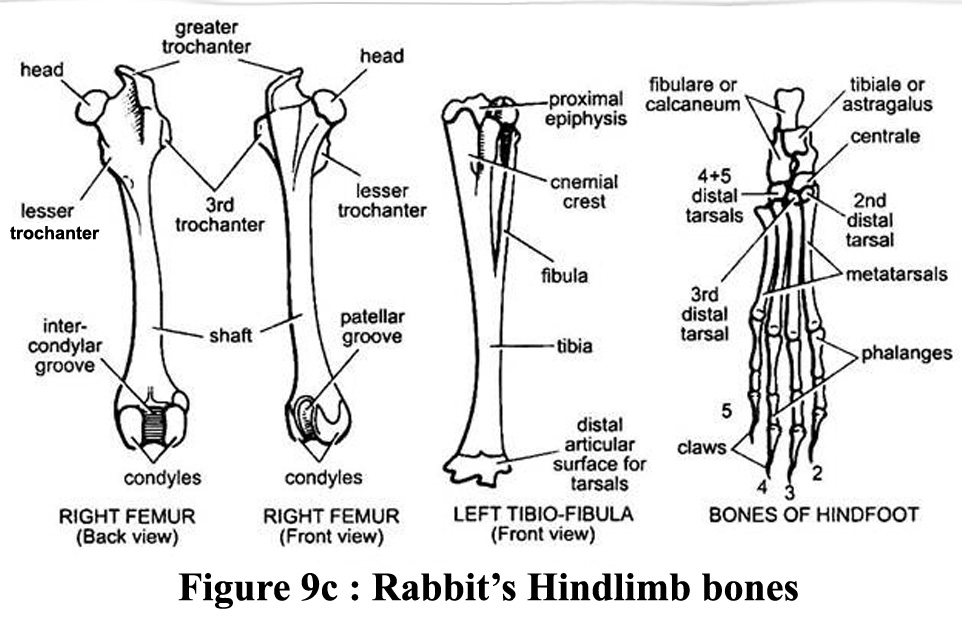
The radius
and the cubitus (also called the ulna, the longest of the two foreleg
bones) in the anterior limb are in contact but not fused (Barone et al.,
1973). However, in the posterior limb, the tibia and the perone are completely
fused in the distal part. The "remaining" part of the perone
is called the fibula. Illustation of bones of the 2 legs is available
in figures 9b and 9c |
| |
|
|
|
|
|
|
Figure
9a : Vertebrae of a rabbit
|
Figure
9b and 9c : Bones of the forelimb and of the hindlimb of a rabbit
|
|
| |
|
|
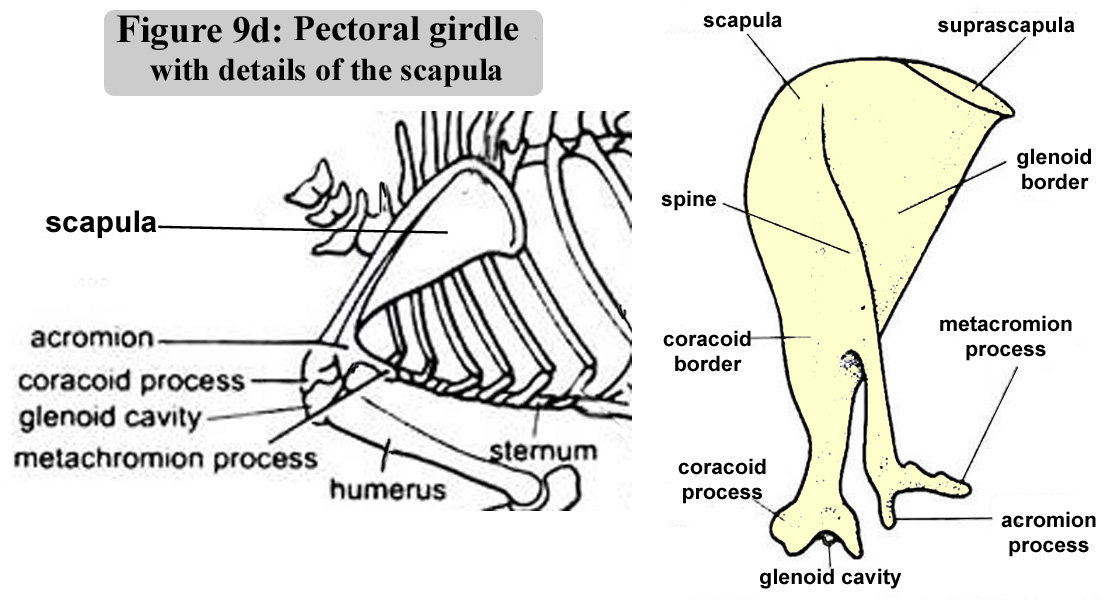
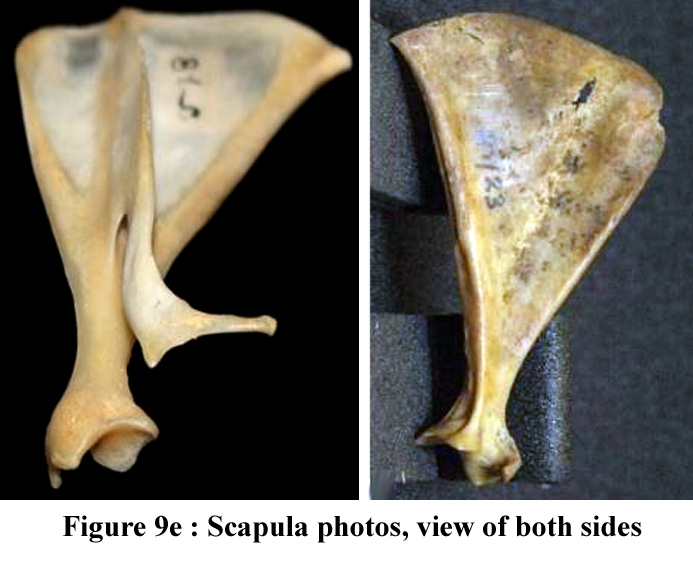
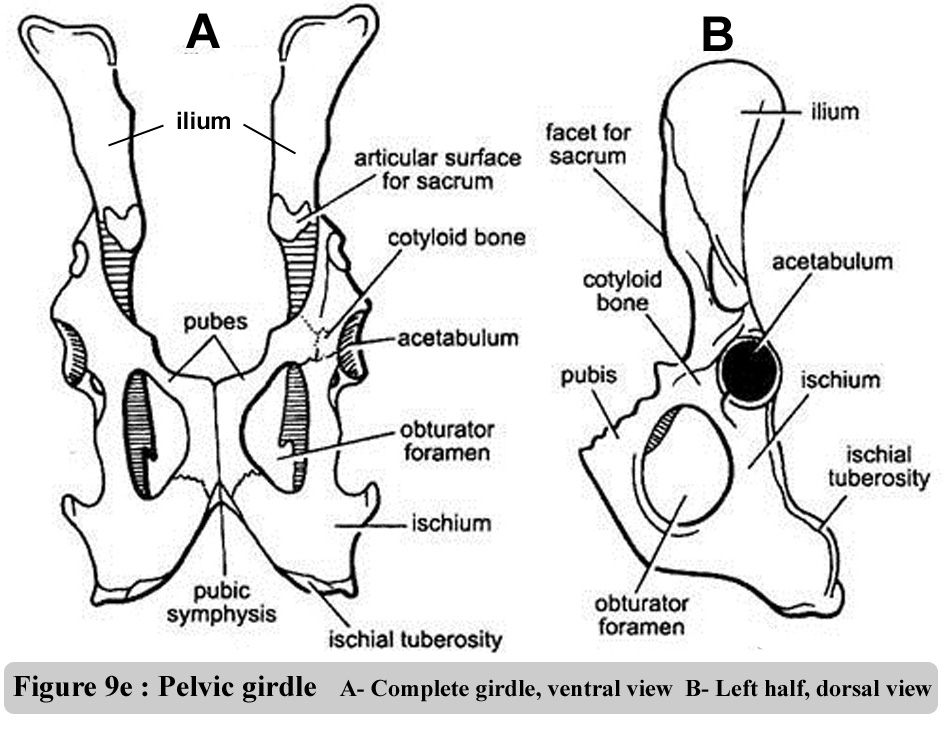
The oganisation
of pectotal and of the pelvic grids are illustrated in figure 9d and 9f.
The figure 9e contains photos of the scapula (pectoral gridl) |
| |
|
|
|
|
|
|
|
Figure
9d : Pectoral girdle with details of the scapula
|
Figure
9e ; Photos of the scapula (both sides)
|
Figure
9f : Pelvic girdle - Ventral & ½ dorsal views
|
|
| |
General
bone structure and bone growth |
| |
|
|
B0nes
have two different structures. On the periphery there is a hard dense
"layer", which is the compact bone. The middle part consisting
of small cavities is the spongy bone whether compact or spongy, adult
bone tissue is laminar. The structure of the compact bones rests on a
beautiful order of concentric laminae. The laminae in the spongy bones
form an irregular pattern of fine rows. The bone marrow inside the bones
is responsible for producing components in the blood and immune system,
that is, red blood cells, white blood cells and platelets. |
| |
|
|
|
| |
Bone
cells |
| |
|
|
Three
types of cell are involved in the life and metabolism of the bone tissue.
These are the osteocytes, osteoblasts and osteoclasts, fed by a network
of blood vessels.
The osteocytes
are cells trapped in the bone matrix but which communicate with each
other by fine extensions connected together. These cells are "mechano-receptors"
sensitive to pressure variations exerted on the bone matrix.
The osteoblasts
are found on the surface of the trabecular bone and they synthesize
the elements of the bone matrix, as well as facilitating their calcification.
They communicate with each other and with the matrix osteocytes, via
a system of specialised cellular unions.
The osteoclasts
are cells with several nuclei. They locally reabsorb the surface of
the bone trabeculae in the form of small erosions, which are the origin
of an initial reabsorption area. They are not related to the other bone
cells by unions but are paired by soluble mediators that move from one
cell to another.
|
| |
Skeleton
growth |
| |
|
|
Initially,
the development of the skeleton is ensured by points of ossification
(from fibrous cartilage membranes). The combined work of the osteoblasts
ensures the growth of the bones in diameter and thickness by continually
renewing the bone mass. The osteoblasts are predominantly active during
the growth period. The conjugation cartilage at the base of the epiphysis
of each bone (or epiphyseal plate) is responsible for growth in length.
After 140-150 days, growth in length stops, the length of the rabbit stabilizes
and the epiphyseal plate "closes", that is, the conjugation
cartilage disappears, mainly due to the action of steroid hormones (Gilsanz
et al., 1988). Average skeletal growth speed is slightly slower than that
of overall body mass from birth to 5-6 weeks of age : allometric coeficient
a = 0.82 (Cantier et al., 1969). After this age, speed of growth of the
skeleton is is about one half of that of the whole body mass :a: 0.55,
and skeleton growth stops when the rabbits are 5-7 months depending of
the breed. |
| |
|
|
|
| |
Muscles
and Musclar growth |
| |
|
|
| Quantitatively
speaking, the most important muscular masses are found in hindquarters.
These are the loins and thighs. The skeletal muscles ensure the
animal's movements and for this reason are attached to the skeleton
in 2 or 3 places, so that movement is generated when they contract.
From the "meat"
point of view, the Longissimus dorsi muscle has the greatest
mass (Swatland, 1994). From the anatomical point of view, it is
a succession of different muscles: the Long dorsal,
Long spinal and the Longissimus lumborum (Cantier et
al., 1974).
Very diverse
in shape, size and function, skeletal muscles are also characterized
by strong tissue heterogeneity.. The nerve tissue (afferent motor
fibres and efferent sensitive fibres) ensures muscular contraction
and relaxation control. The blood vessels guarantee the supply
of nutrients and oxygen and remove products from the cellular
catabolism. The muscle fibres constitute 75 to 90% of the muscle
volume and are the base element of the striate skeletal muscle
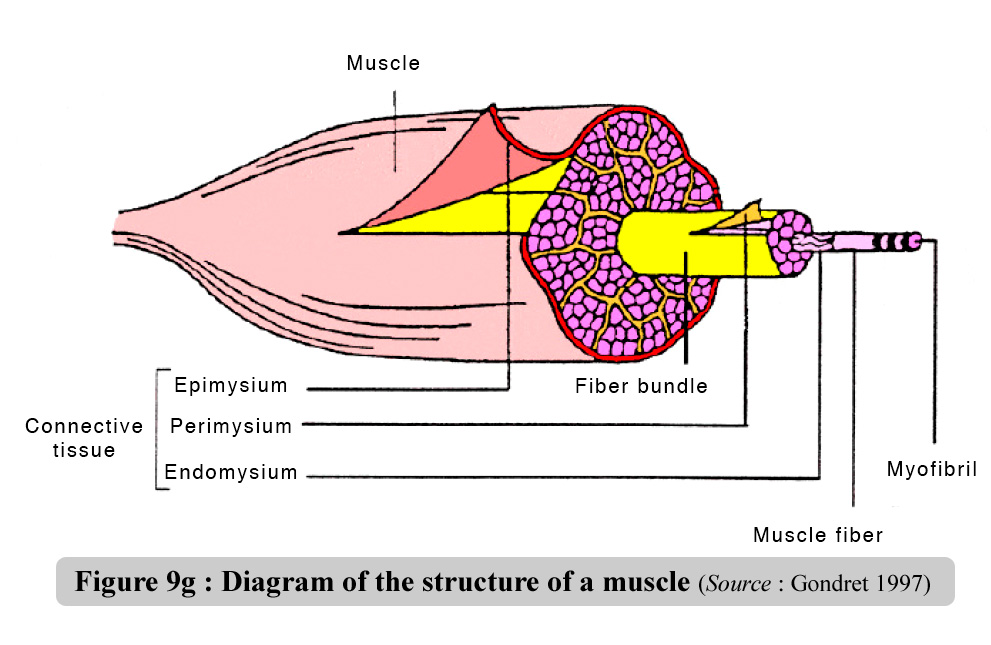
(Gondret, 1997). They are covered in a delicate sheath of conjunctive
tissue called endomysium which is rich in collagen (figure 9g).
|
|
|
Figure
9g : Diagram of the structure of a muscle - Source
: Gondret (1997)
|
|
| |
Muscle
fibers and muscular growth |
| |
|
|
The muscle fibers
are grouped together in primary bundles covered in a fine conjunctive
sheath called perimysium. The adipose cells, called intracellular adipocytes,
are found amongst the primary bundles. The primary muscular bundles
are in turn grouped together into secondary bundles, covered in a thicker
sheath of perimysium. The epimysium encloses all the thick secondary
bundles to form the cover/envelope of the muscle itself. The conjunctive
section at the muscle ends forms the tendons and aponeuroses responsible
for binding the muscle to the skeleton. The areolar tissue between the
interfibrilar spaces forms an intricate network rather like a more or
less developed elastic mesh. Its function is to provide the muscle with
a framework capable of organizing the muscular fibres, packing the nervous
and vascular elements and providing the necessary lubricated surfaces
for the muscular fibres to move amongst each other and amongst the bundles
(Doutreloux, 1992).
From a quantitative
point of view, muscular speed of growth is a bit more rapid than that
of the whole body from birth to 10-12 week of age, according to the
breed or the line : allometric coefficient a = 1.2. After this period
corresponding to the establishment of puberty, muscular growth slows
sharply down and is about one half of that of the whole body : allometric
coefficient a : 0.50 (Cantier et al. 1969). .Muscular growth ends when
the rabbit is 5 to 7 month of age , depending of the breed. However,
individual muscles follow different patterns of longitudinal and cross-sectional
growth, so that their functional capacities (force, range of contraction)
and mutual functional relationships are age-dependent. Fibre type distribution,
fibre cross-sectional area and compactness, colour and metabolic characteristics
varied according to age. The effects of eventual feeding treatment are
low in comparison with age effects.
|
| |
Muscle
fibers and myofibrils
|
| |
|
|
Muscular fibre is
an elongated, tubular, plurinucleate cell. It is the basis of motor
activity Most of the sarcoplasm in the muscular cell is occupied by
contractile elements called myofibrils. There are three different types
of filaments in the myofibrils: thick ones (150 Å in diameter),
which are basically formed by myosin, and fine ones (70 Å in diameter),
formed by actin. They also contain tropomyosin and the troponin complex.
The intermediate filaments constitute a network of longitudinal and
transversal filaments associated to the sarcomere. When the bundles
of fibres go from one end of the muscle to the other, parallel to the
main axis, the muscle is described as "fusiform" (i.e.,
the Biceps femoris). When they form a more or less definable
angle, in relation to the main axis, the muscle is "penniform"
(from penna, feather); an example of this is the Longissimus lumborum.
|
| |
|
|
Myosin and
actin have a key function in the architecture and a key enzymatic
function in muscular contraction, during which chemical energy is transformed
into mechanical energy. Muscular contraction is produced when the fine
actin filaments slip between the thick myosin ones. The energy required
for contraction comes from ATP hydrolysis via the myofibrillar ATPase.
The ATP is then regenerated by the aerobic or anaerobic metabolism.
Depending on the speed at which the fibres contract in the centre of
a muscle and on the major type of metabolism working to regenerate the
ATP used in the contraction, different types of fibres are differentiated;
their functional characteristics are listed in table 3. Bearing in mind
the very variable myoglobin content of one type of fibre or another
and the fact that some muscles are formed exclusively by one type of
fibre, the rabbit's muscles are either red or completely white
|
| |
|
|
Table
3. Characteristics of the different types of muscular fibres.
Source: Gondret (1997).
|
| |
|
|
|
Characters
|
Types
of Fibres
|
|
I
|
IIA
|
IIX
|
IIB
|
|
Contraction
sped
|
slow
|
fast
|
fast
|
fast
|
|
Fatigue
resistance
|
+++
|
++
|
?
|
?
|
|
|
|
Motor
plate surface
|
+
|
+++
|
?
|
+++
|
|
Sarcoplasmic
Reticulum
|
+++
|
+++
|
?
|
+++
|
|
Transversal
tubules
|
+
|
+++
|
?
|
+++
|
|
Color
|
red
|
red
|
white
|
white
|
|
Myoglobin
|
+++
|
+++
|
++
|
+
|
|
Capillairy
density
|
+++
|
++
|
?
|
+
|
|
Number
of mitochondrion
|
+++
|
+++
|
++
|
+
|
|
Collagen
content
|
+++
|
++
|
?
|
++
|
|
Section
area
|
+
|
++
|
+++
|
++++
|
|
|
|
Use
of glycogen
|
+
|
++
|
++
|
++++
|
|
Use
of lipids
|
+++
|
+++
|
?
|
+
|
|
|
|
Myofibrillar
ATPase
|
+++
|
++
|
?
|
+
|
|
Hexokinase
|
+++
|
++
|
?
|
+
|
|
Phosphorylase
|
+
|
++
|
?
|
++
|
|
Anaerobic
enzymes of glycolyse
|
+
|
++
|
++
|
++
|
|
Oxidative
Enzymes (aérobies)
|
+++
|
++
|
++
|
+
|
|
| |
Repartition
of main muscles on the rabbit carcass |
| |
|
|
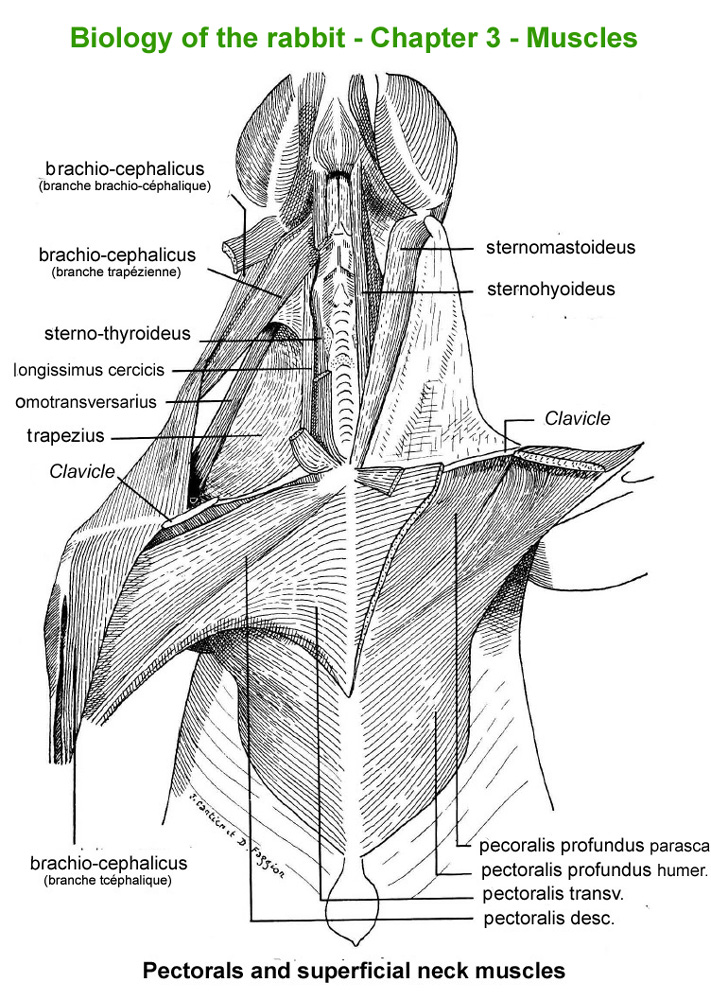
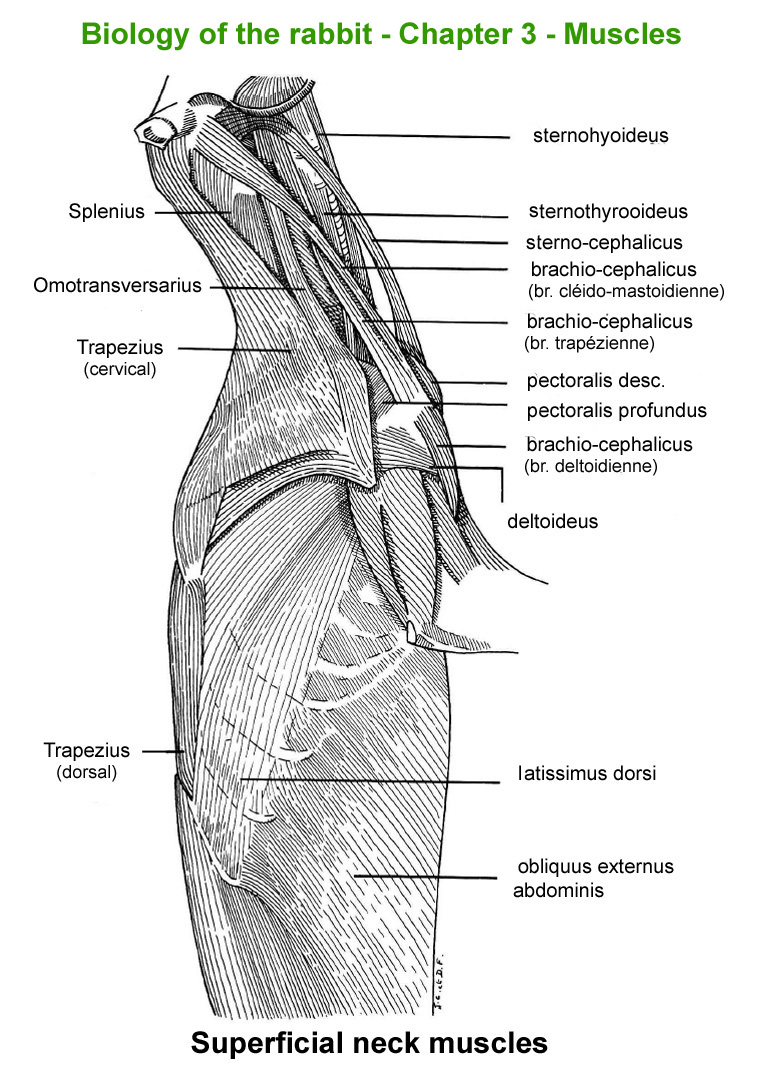
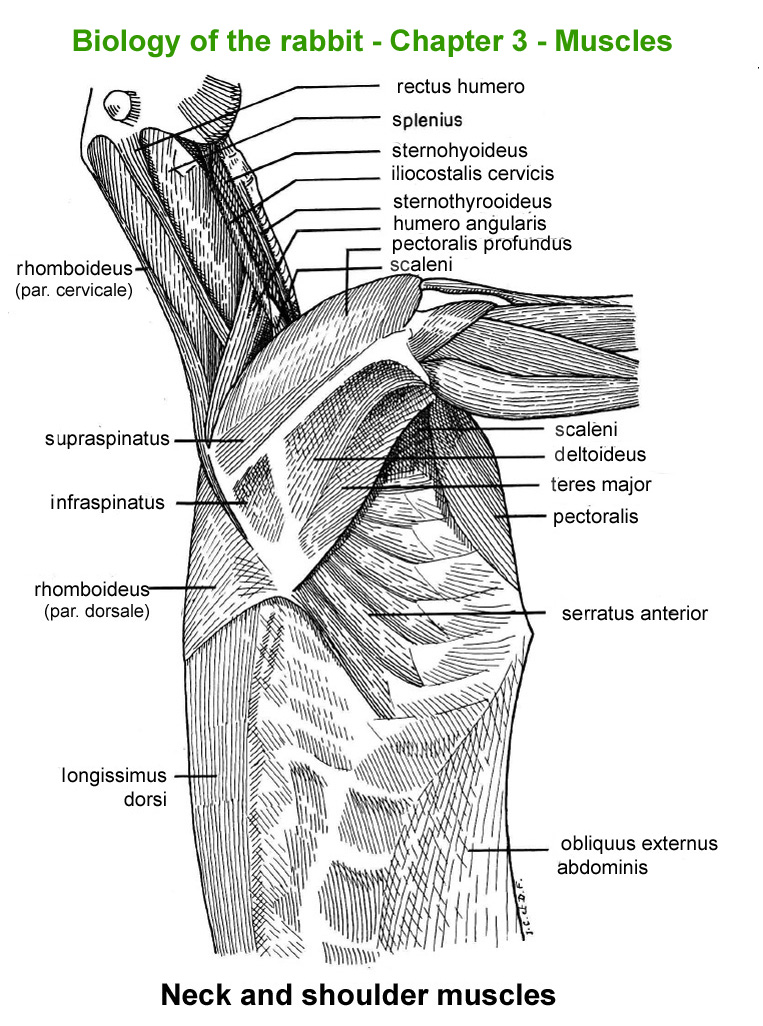
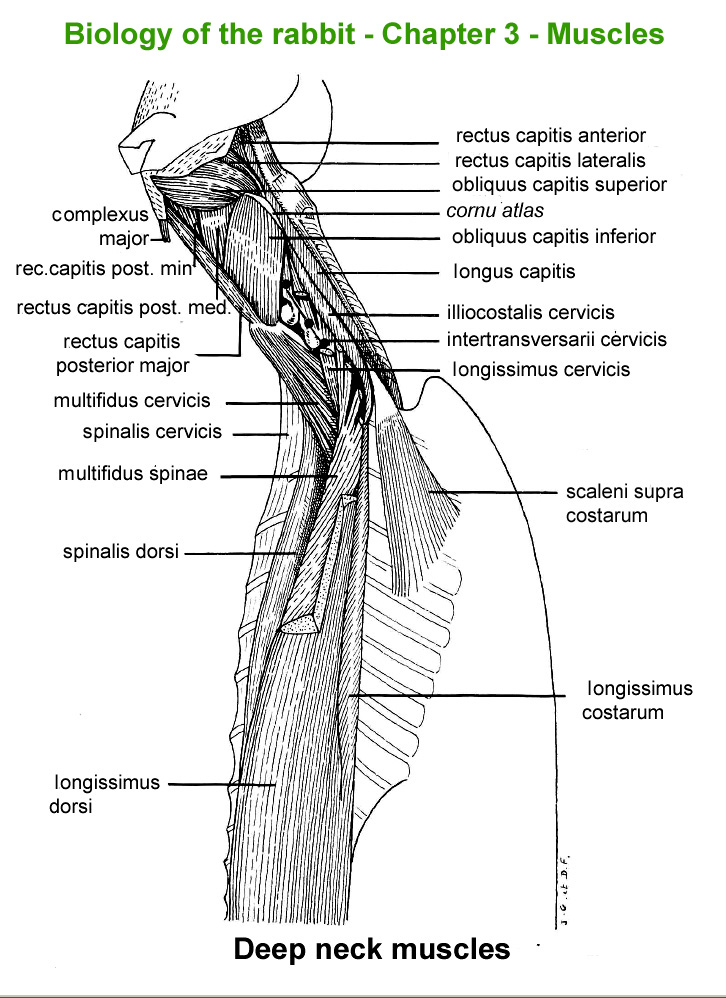
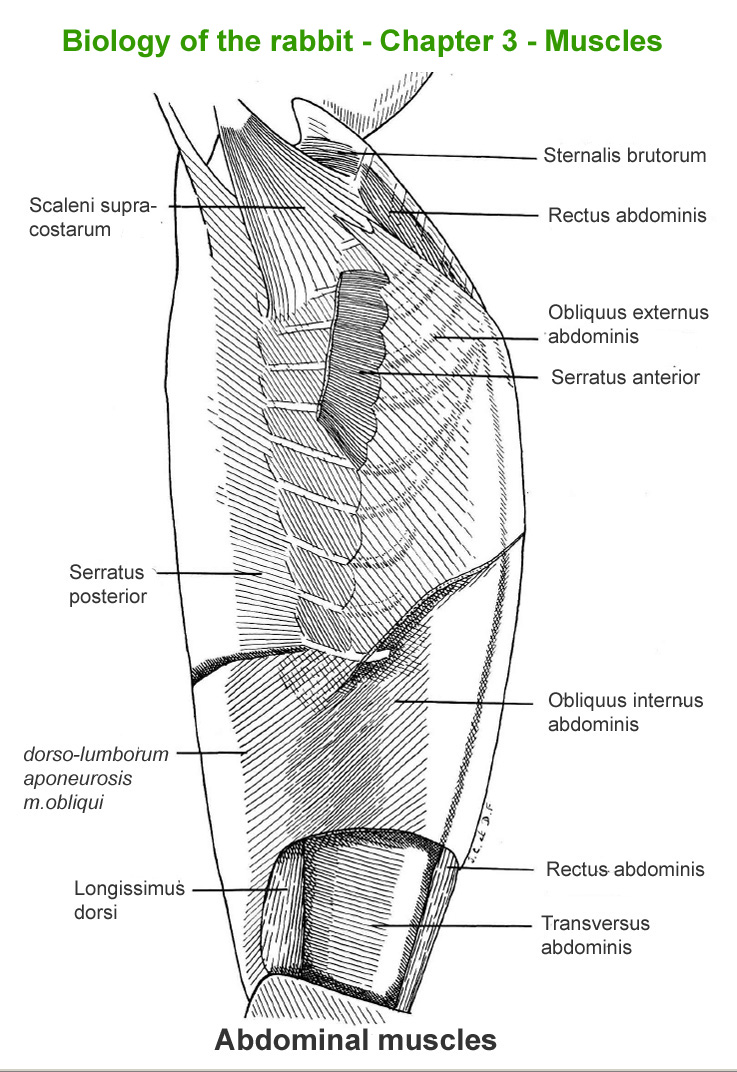
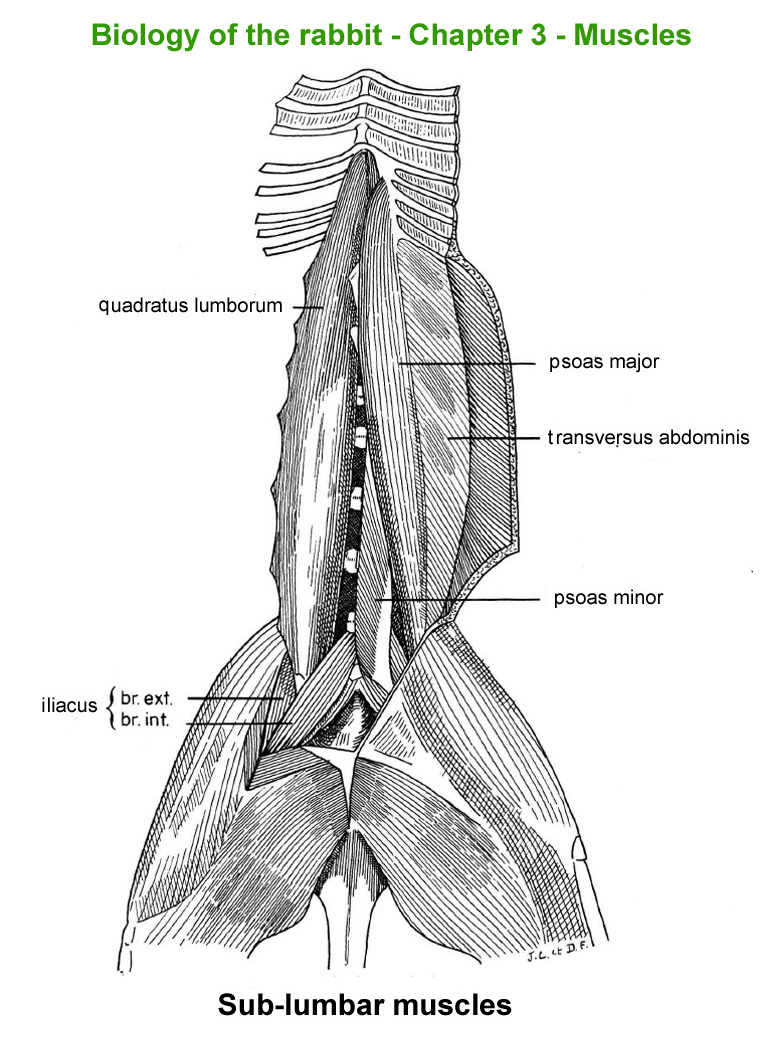
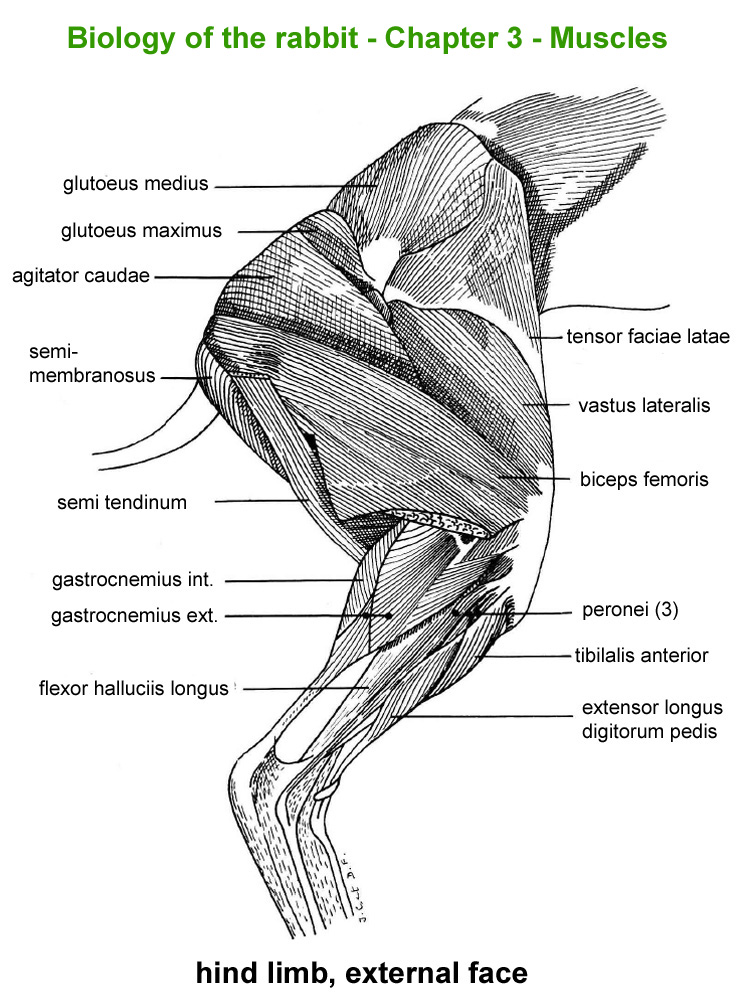
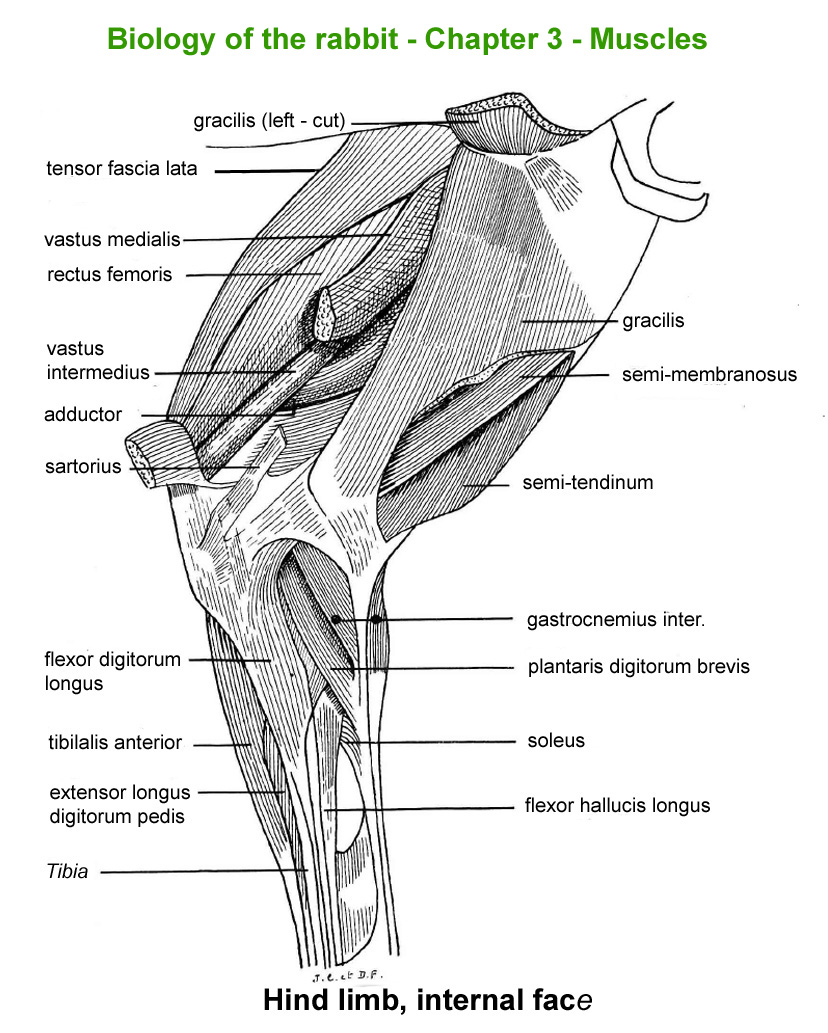
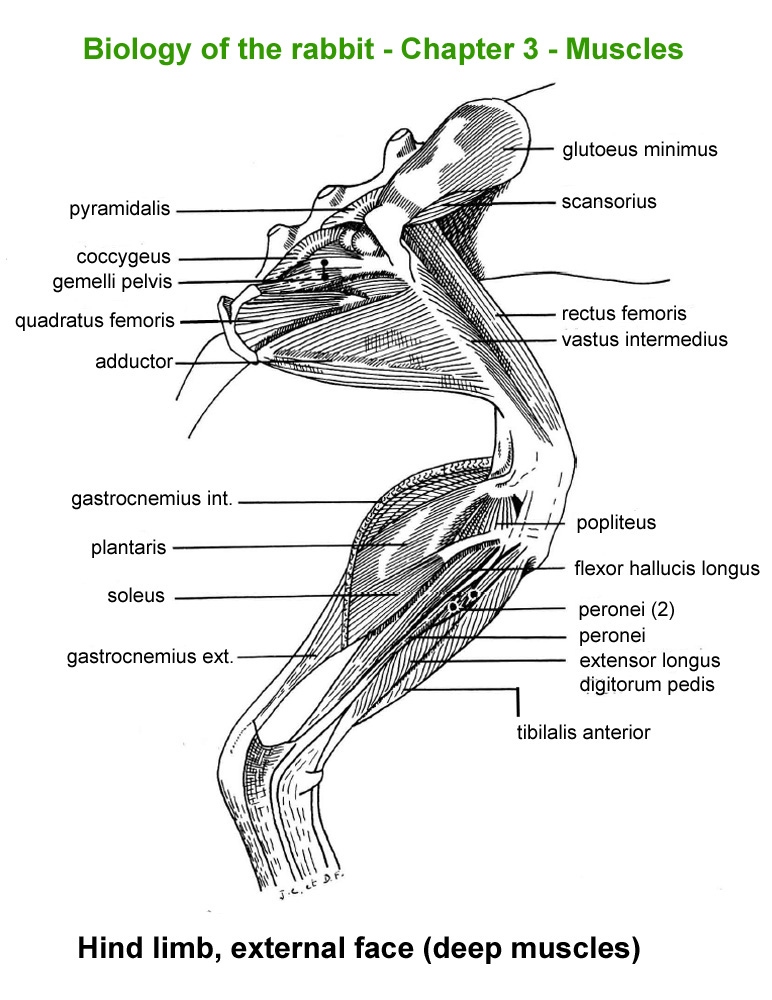
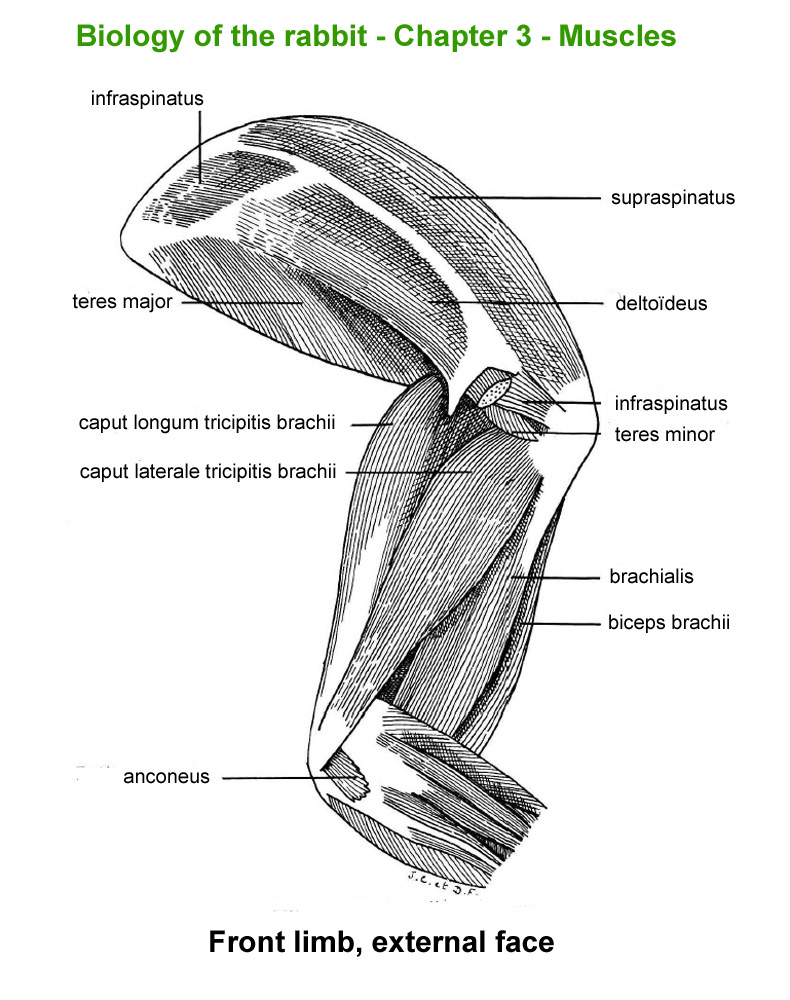
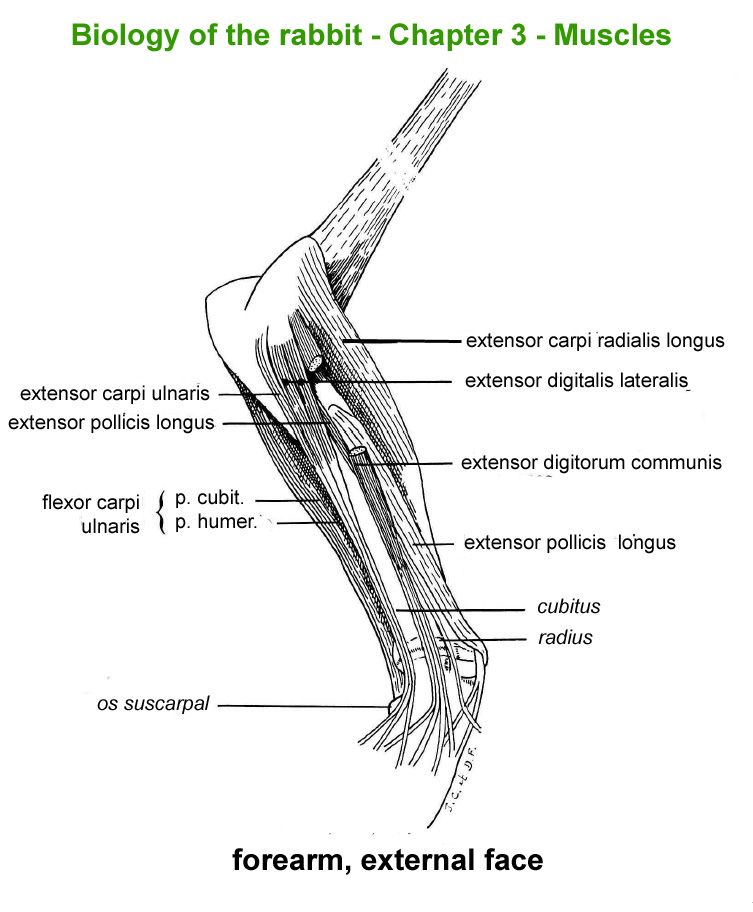
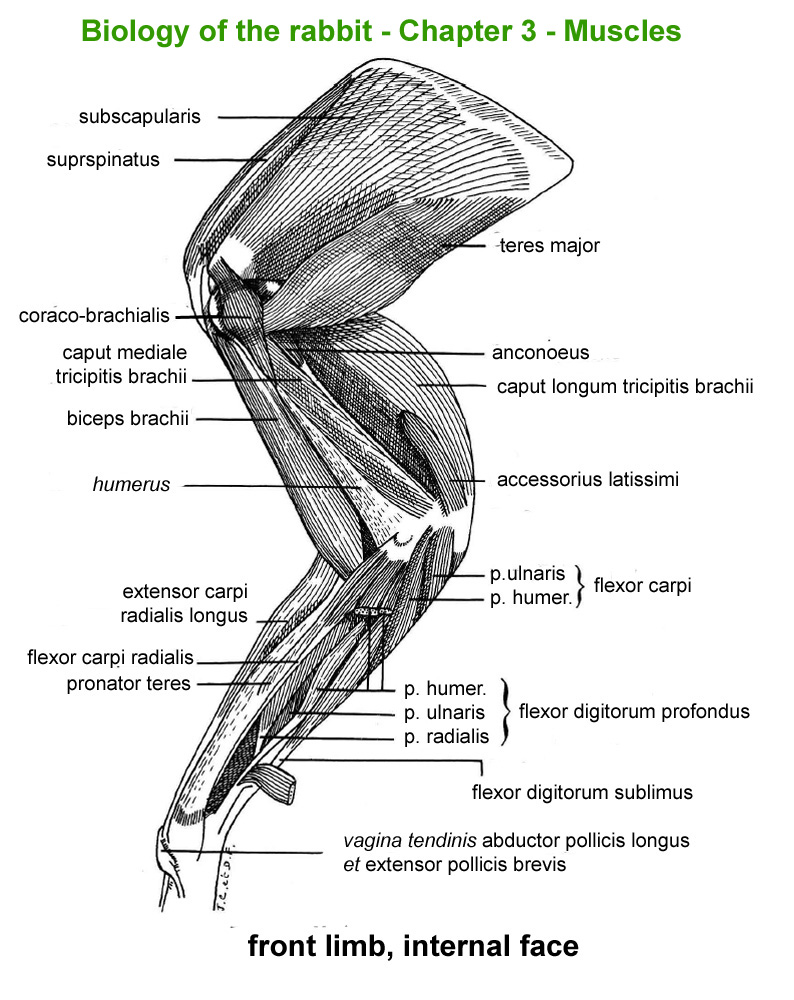
The
main skeletal muscles observable during a dissection are described in
figure 9h. For more details it is possible to refer to the original article
published in French in 1968 by Cantier
and Vezinhet with explanations for each muscle. In this article, for
each cited muscle, the French name and the international Latin name are
given |
| |
|
|
|
Figure
9h : Main skeletal muscles
|
|
|
|
|
|
|
|
|
Pectoral
and superficial neck muscles
|
Superficial
neck muscles
|
Neck
and shoulder muscles
|
Deep
nek muscles
|
Abdominal
muscles
|
Sub-lumbar
muscles
|
|
|
|
|
|
|
|
|
Hind
limb, external face
|
Hinf
limb, internal face
|
Hind
limb, external face (deep
muscles)
|
Front
limb, externam face
|
Forearm,
external face
|
front
limb, internal face
|
|
| |
Adipose
tissue |
| |
|
|
At slaughter
age (10-12 weeks), the dissectable adipose tissue, 1.4-1.7% rabbit's live
weight, is composed by abdominal (peri-renal) fat (53%), scapular fat
(41%) and inguinal fat (6%) . In addition to these organised tissues the
presence of small quantities intermuscular adipose tissue (fat nodules)
and some intramuscular adipose cells must be mentioned. In the rabbit
like in some other species adipose tissue is one location of lipids synthesis
(in addition to liver) and of lipiolyse . It acts as a reserve tissue
which develops especially once that metabolic requirements of the growth
of noble tissues (nervous, skeleton, muscles) have been reached. If in
a slaughter rabbit (10-11 weeks) , dissectable adipose tissue represents
about 1.3 to 2% of live weight), in some particulaly fat adults, abdominal
fat alone can represent up to 25% of live weight or more. |
| |
|
|
From
birth till 5-6 weeks of age, weight development of adipose tissue is slower
than that of the whole body mass : allometric coefficient a= 0.82. After
this age adipose tissue begins to grow faster than the whole body (a=
1.87) till about 10-12 weeks of age At this period of rabbit’s life,
skeleton and muscular tissue developments begin both to slow down and
adipose tissue begins to grow very rapidly : a=3.21 till adulthood (Cantier
et al. 1969) |
| |
|
|
The
presence of intermuscular adipose tissue inside of the different carcass
cuts (fat nodules), and the simultaneous presence of adipose cells inside
of muscles are largely responsible of juiciness and of the taste of rabbit
meat. The importance and lipid composition of these internal adipose components
are sharply correlated with the more visible abdominal fat. For this reason,
quality of a rabbit carcass is partly estimated by the importance of visible
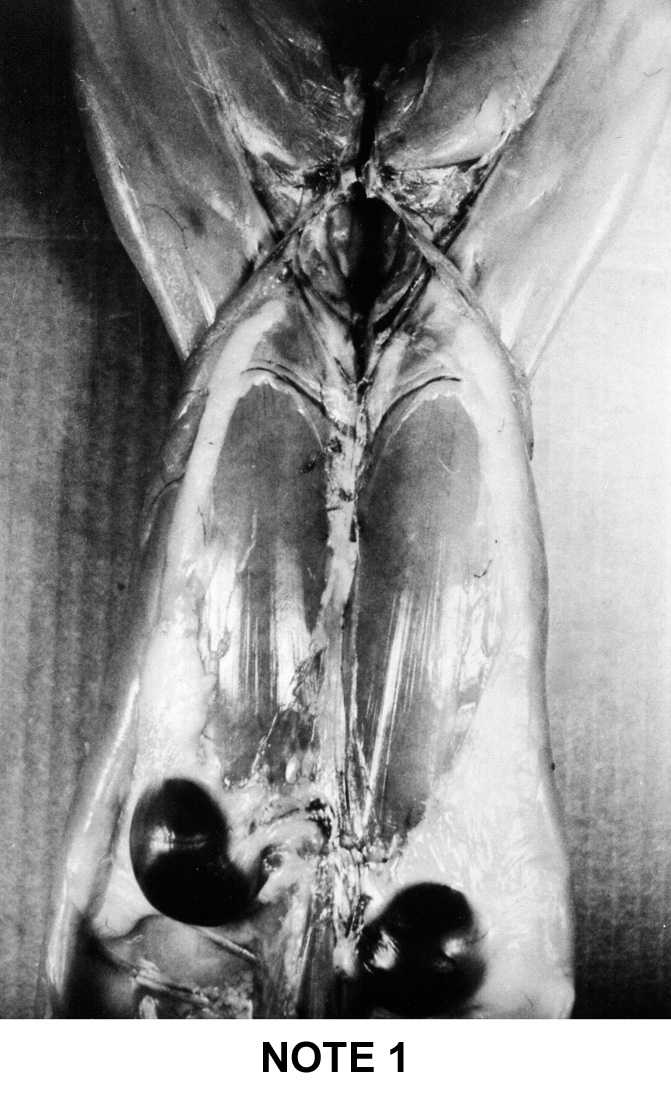
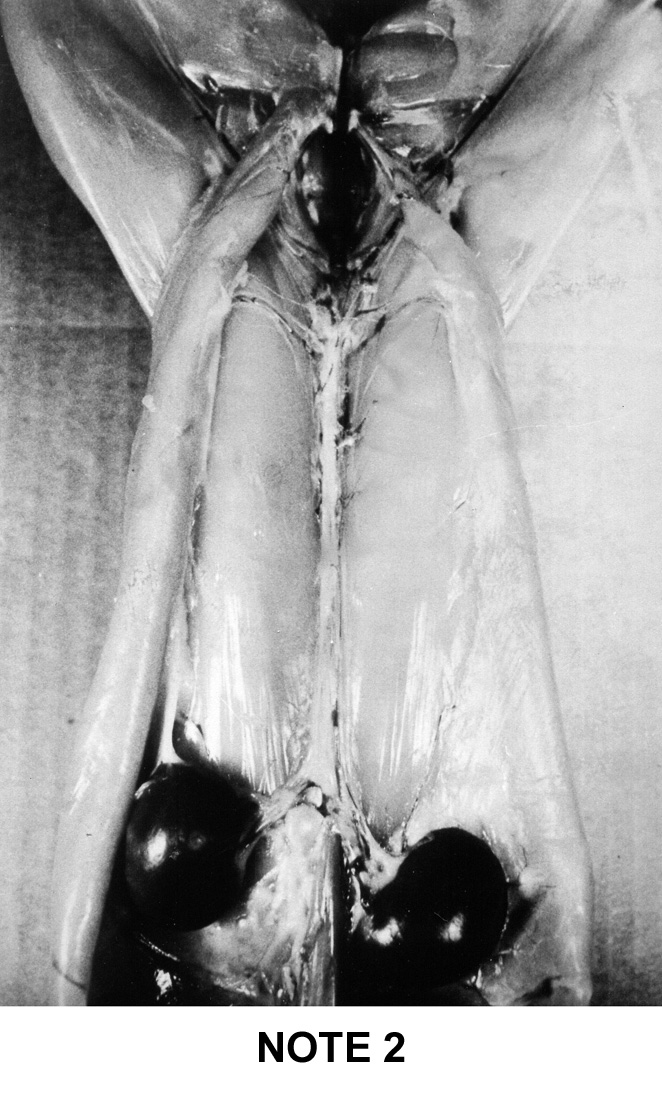
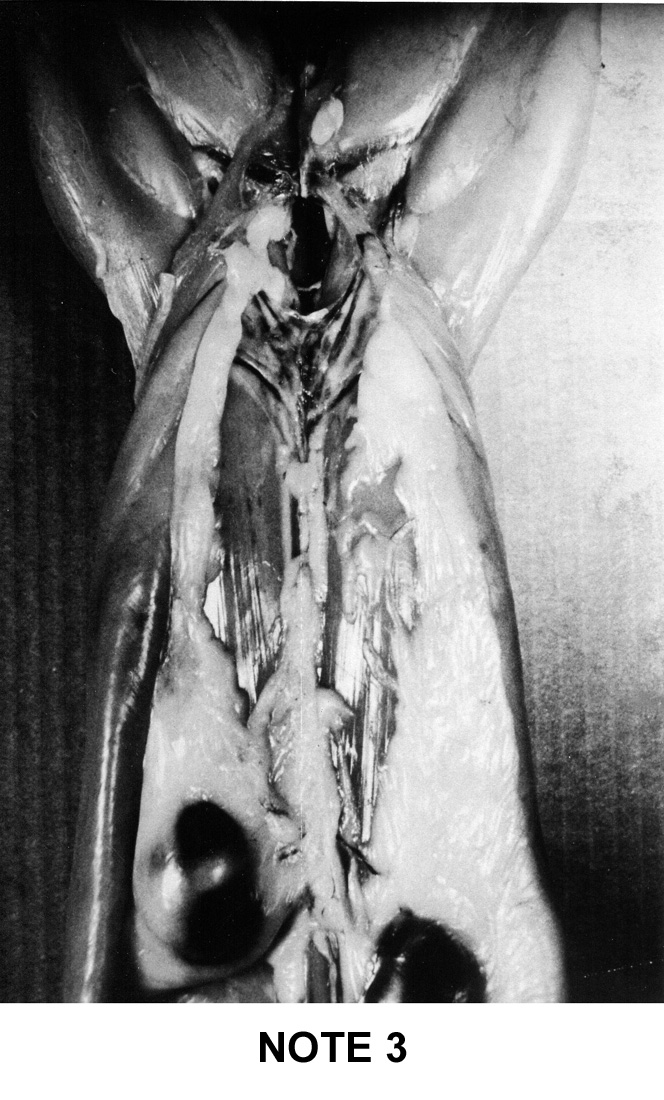
abdominal fat . To help the estimation of the general adiposity of a carcass
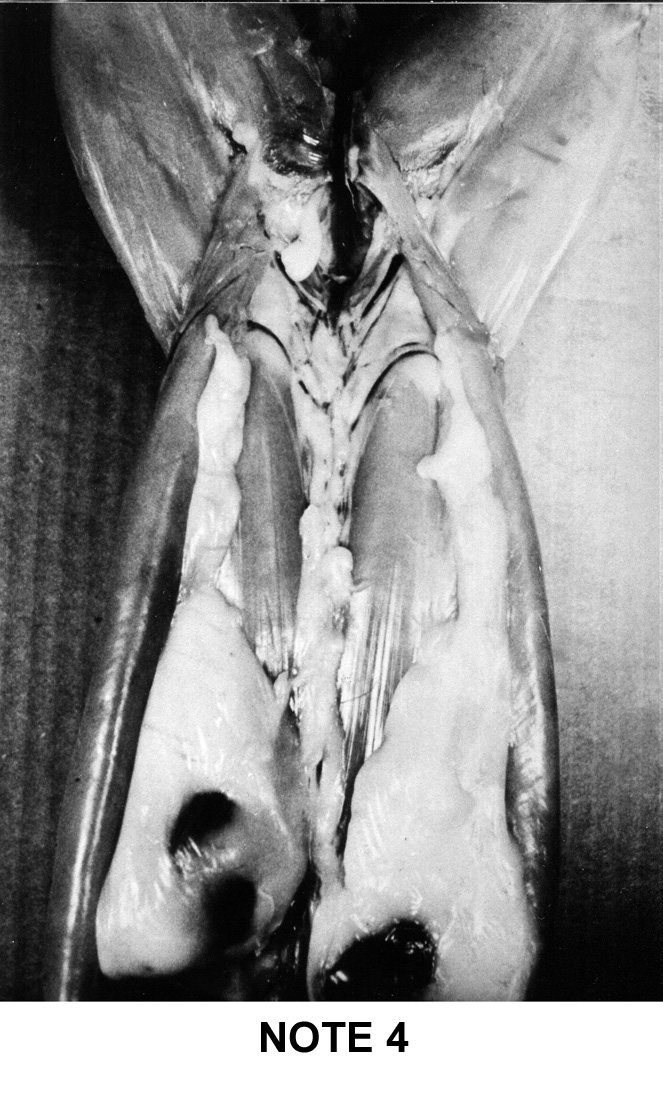
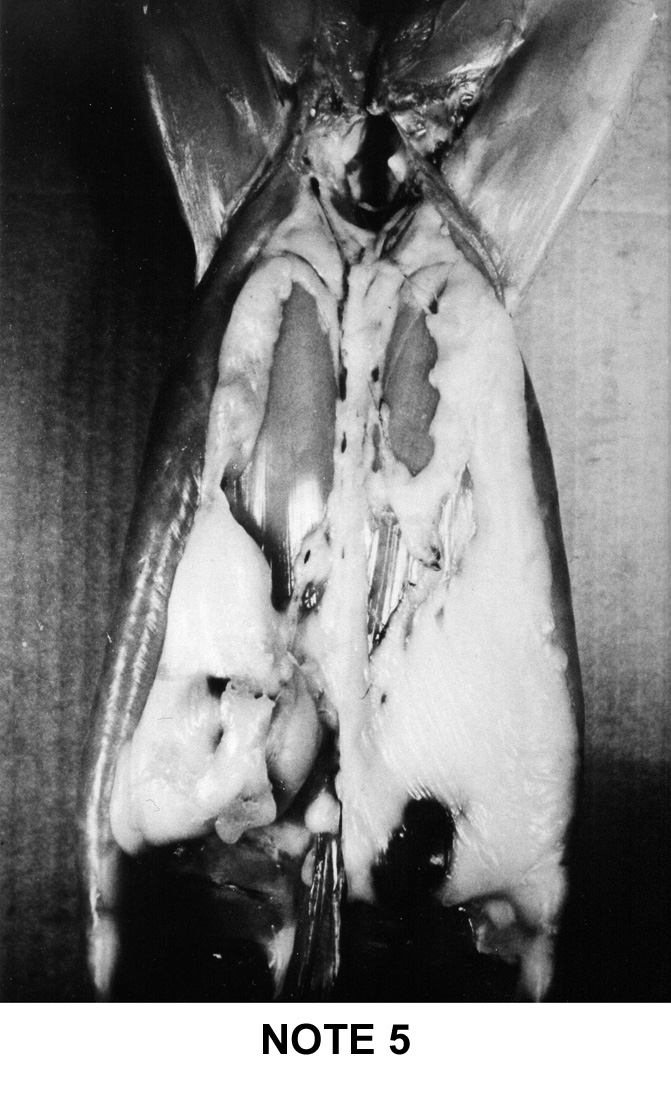
a simple system of visual notation was established in France: note 1 for
the too lean carcasses up to note 5 for the too fat carcasses. This system
is described below on figure 9i |
| |
|
|
|
Figure
9i : French system of notation of rabbit's carcass adipodity
|
|
Note
1
|
Note
2
|
Note
3
|
Note
4
|
Note
5
|
|
|
|
|
|
|
- No or
little fat
- The
2 kidneys are uncovered and protruding
|
- Two
individualized lateral fat masses reaching the groin in two
narrow bands.
- Absence
of fatty cord along the spine
- The
right kidney is lightly covered on the outer side
- The
left kidney adheres to fatty tissue but is not at all covered
|
-
Two fairly large lateral fat masses over the
entire length abdominal cavity and the presence of a thin
fatty cord along the spine.
-
The right kidney is half covered by adipose
tissue
-
The left kidney is lightly covered, its outline
remains visible
|
-
Two thick and wide fat masses running the
length of the abdo-minal cavity, and the presence of a very
distinct fatty cord along the spine.
-
The right kidney is two-thirds coated, its
upper surface remains visible.
-
The left kidney is covered over more than
half of its surface
|
- Two thick and wide fat masses encompassing the fatty cord
central in the renal area and covering more than half of the
saddle surface.
- The right kidney remains slightly visible.
- The left kidney is completely covered.
|
|
| |
Special
case of the newborn |
| |
|
|
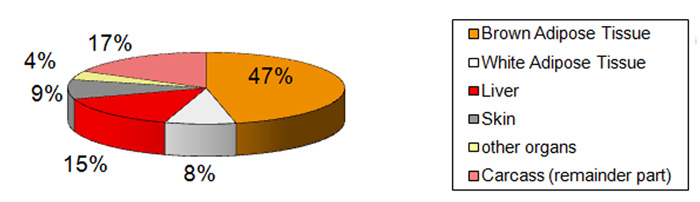
At
birth the newborn rabbit contains about 5.8% of lipids, wile later
this proportion varies from 10% to 15% according the age or reproduction
stage. But if the lipids content is clearly lower, the main diffreence
is in the repartion in the young rabbits of fat tissues (energy reserves).
The main lipids reserve is in the brown adipose tissue and partily in
the liver .Lipids of the white adipose tissue represents only 8% ot the
total (figure 09j) . The role of these particular tissues (brown and white
adipose tissue) is explained the section 7.4 devoted to the newborn biology
. But in few words the brown adipose tissue is a lipids reserve exclusively
used for thermoregulation under nervous control. This tissue will disappear
some weeks after weaning (modification of type of metabolism). Lipids
of all other tissues participe normally in the general metabolism. |
|
Figure
09j : Repartition of lipids of a newborn rabbit in the different
tissues and organs
(according to Dawkins et al., 1964)
|
|
|
|
| |
|
|
|
| |

Biology
of the Rabbit -
Index of the Website
|
| |
| |
| |