| |
4.1
Some anatomy reminders : situation in adult |
| |
|
|
The
alimentary canal of an adult (4-4.5 kg) or subadult (2.5-3 kg) rabbit
is about 4.5 to 5 metres long. Its position in the abdominal cavity is
shown in figure 10. The digestive segments and their main characteristics
are described in figure 11 |
| |
|
|
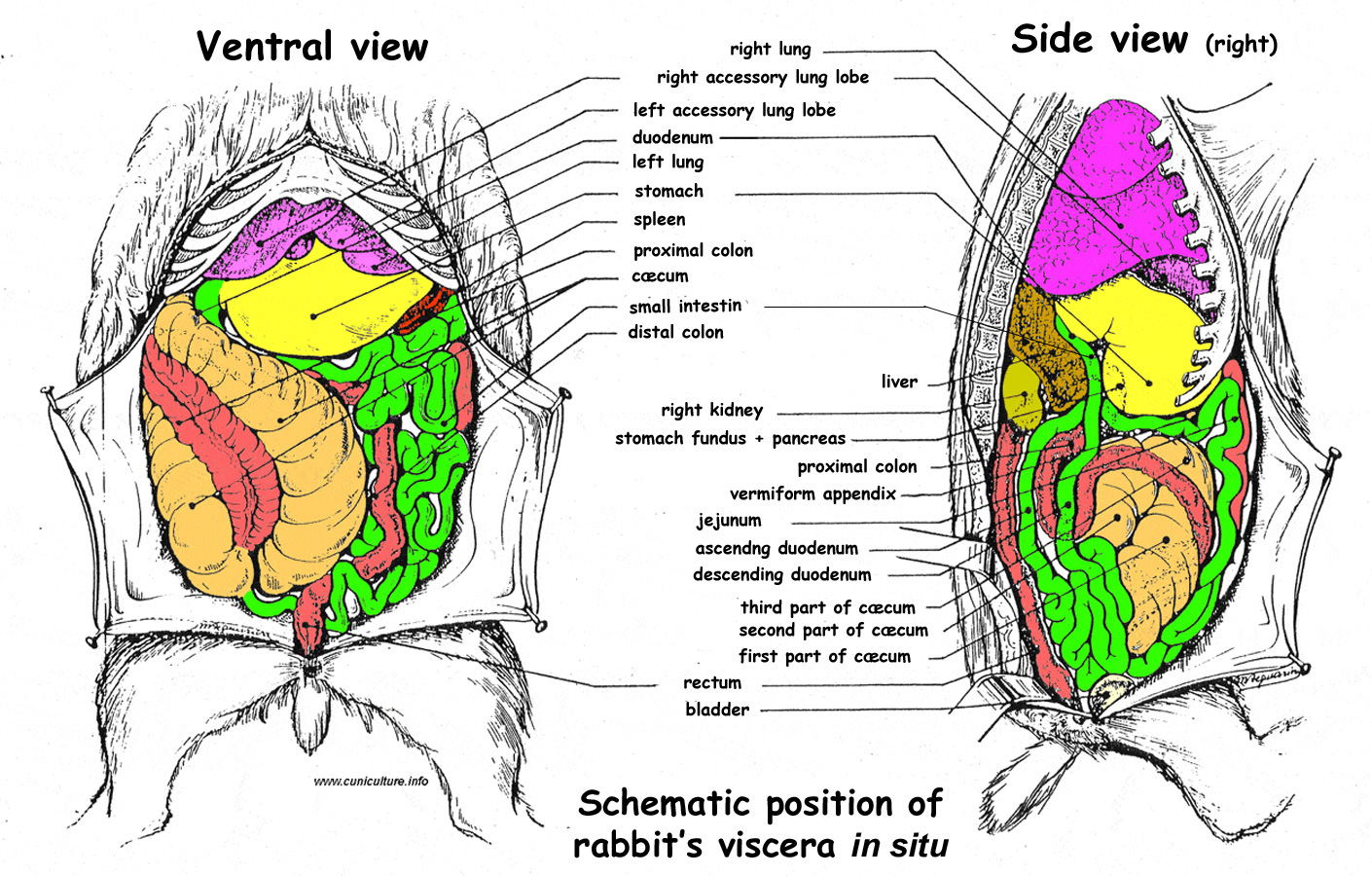
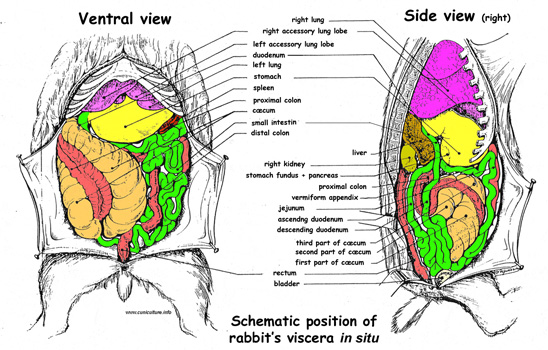 Figure 10 : .
Position of the rabbit's viscera in the abdomen
Figure 10 : .
Position of the rabbit's viscera in the abdomen . Source:
Domini (1967). |
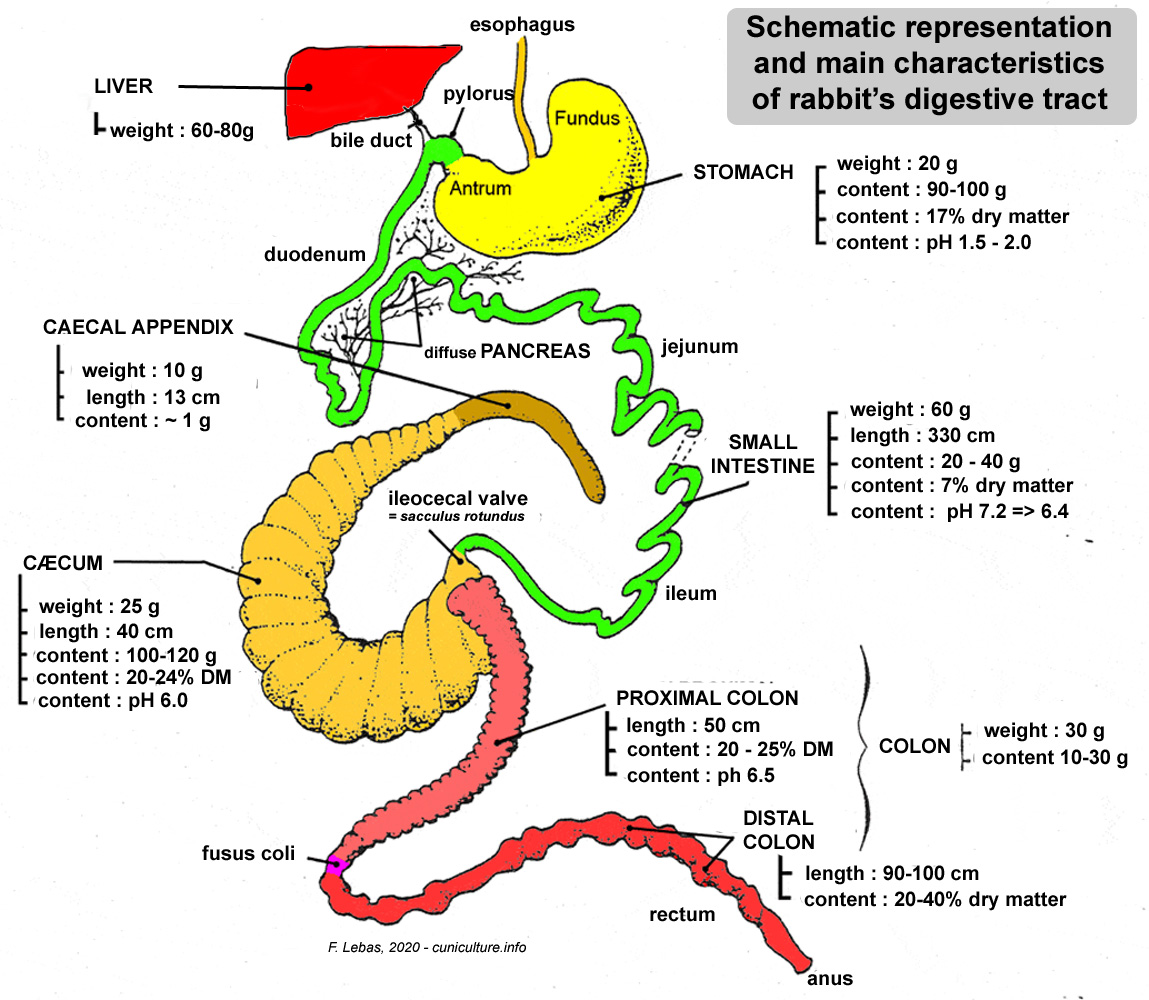
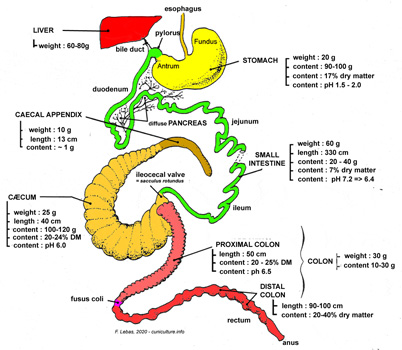 Figure
11. Diagram of the different organs in the digestive tract.
Source: Lebas et al.
(1996a).
Figure
11. Diagram of the different organs in the digestive tract.
Source: Lebas et al.
(1996a). |
| |
|
|
|
| |
Mouth |
| |
|
|
As
previously indicated (chaper 3), the teeth grow continuously and their
masticatory function is moderate. The salivary glands (parotid, maxillary,
sublingual and zygomatic or orbital) produce saliva containing very little
amylase (25 µmol of maltose obtained from the starch per mg of salivary
protein, compared to 250 or 450 for the pancreatic juice). The amylase
content is independent of the starch content of the feed ration or whether
the animal has eaten or not (Blas et al., 1988). |
| |
Esophagus |
| |
|
|
The
oesophagus is between the trachea and the spinal column. It allows the
alimentary bolus to move from the mouth towards the stomach only. Regurgitation
never occurs, even by accident |
| |
Stomach |
| |
|
|
|
The stomach
is an elongated sac with a mucous lining. The oesophagus is connected
to the stomach by the cardia. The "blind" part of the
stomach is the fundus (fundus in Latin) and the opposite
region is the pyloric antrum (antrum pyloricum), which
ends in the pylorus. It is endowed with a powerful sphincter that
regulates the transfer of food towards the small intestine.
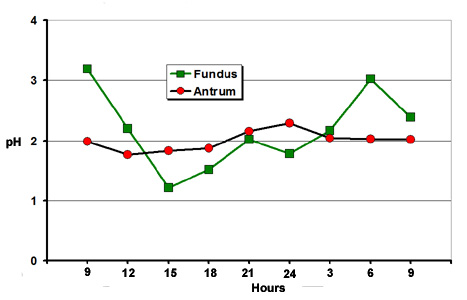
The wall mainly
secretes hydrochloric acid, pepsin and minerals (Ca, K,
Mg and Na). The pH is always very acid but varies noticeably during
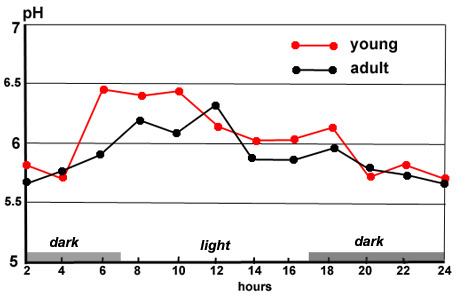
the day, especially in the fundic region (figure 12) The highest
pH (the least acid) is observed when there are caecotrophs (pH
3.5) whilst the most common pH is between 1.5 and 2.0. The secretion
of pepsin and electrolyte varies in the same way and depends,
in particular, on the ingestion rate of caecotrophs (Beauville
and Raynaud, 1964; Salse et al., 1982). The stomach content of
a 9-week-old rabbit is 90-120 g of fresh matter, depending on
the time. The dry matter content varies between 16 and 21 % (Gidenne
and Lebas, 1984).
The stomach
wall of a very young kit (1 week old) secretes pepsin,
the optimum pH àf which is between 1.8 and 2.4, as well
as another peptidase, the rennin or chymosin (optimum pH is between
3.4 and 3.8). From 21 days on, the optimum pepsin pH drops to
1.2-1.8 whilst rennin is not found in a 45-day-old or 60-day-old
rabbit (Angelo and Srivastava, 1979).
During lactation,
endopeptidase is responsible for milk coagulation in the stomach
because break of the chain of kappa-casein Pepsin secretion is
important only from the age about of 30 days. In contrast, the
gastric lipase secreted by a small part of the stomach wall around
the cardia reaches a maximum when the kit is 30 days old. It then
decreases rapidly between the ages of 30 and 60 days and until
180 days. It cannot be observed in adults. (Bernadac et al., 1991).
|
|
|
Figure
12. Variations of stomach pH in two regions, depending
on the time of observation. Source:
Gidenne and Lebas (1984)
|
|
| |
Small
intestine |
| |
|
|
The
small intestine which follows the pylorus, measures about 3 m in length
with a diameter of about 0.8 to 1 centimeter. It is conventionally divided
into duodenum, jejunum and ileum, the terminal part. The bile duct which
brings bile from the liver, opens at the beginning of the duodenum, immediately
after the pylorus. Its opening into the duodenum is regulated by the sphincter
of Oddi. Remember that in rabbits bile is secreted almost continuously
by the liver, then stored in the gallbladder before it is evacuated in
the duedenum. The pancreatic duct (also known as Santorini
duct) opens towards the end of the duodenum about 40 cm from the
pylorus. On the wall of small intestine, plaques of lymphoid tissue about
1 to 2 cm in diameter are observed in place. These are Peyer's patches.
The multiple glands present in the wall of the small intestine secrete
numerous enzymes which complement those secreted by the pancreas. |
| |
|
|
The
content of the small intestine is liquid, especially in the first part
and completely empty segments of approximately 10 centimetres are quite
normal. The pH, which is slightly alkaline in the first part (pH 7.2-7.5),
becomes progressively more acid until it reaches 6.2 - 6.5 at the end
of the ileum. |
| |
Caecum |
| |
|
|
|
|
|
Figure
13: Nycthemeral evolution of caecal pH in young rabbits of 5 weeks
and in adult subjects (18 weeks).
Source: Bellier et al., 1995. (Ad libituù
feeding - ingestion of caecotrophs observed from 4h to 12 h. in
young rabbits, and from 8h to14h in adults)
|
The small intestine
ends at the base of the caecum via the sacculus rotondus, which contains
the ileocaecal valve.The wall of the later is formed by lymphoid tissue.The
caecum forms a second reservoir ( the first is the stomach)
and is 40-45 cm in length, with a mean diameter of 3 to 4 centimetres.
It contains 100 - 120 g of a homogenous paste, with a mean of 22 % dry
matter (DM) and a pH close to 6 (figure 13). The caecal wall is invaginated
in the form of a spiral with 22-25 coils, thus increasing the mucosal
surface area in contact with the caecal content. The caecal appendix
(10-12 cm) is situated at the distal end of the caecum and has a clearly
smaller diameter. Its wall is formed by lymphoid tissue.
Very close to the
end of the small intestine or the "entrance" to the caecum,
is the beginning of the colon, also known as the "exit" .
The caecum thus resembles a no exit diverticulum, on the axis small
intestine - colon (figure 11 above). Physiological studies show that
this diverticulum-reservoir is a place of obliged transit; the contents
circulate from the base to the tip, passing through the centre of the
caecum and returning to the base, along the wall.
|
| |
Colon |
| |
|
|
After
the cecum, is the colon of about 1.5 m. It is first characterized by the
presence of haustra (small pocket-shaped bulges) over about 50 cm: this
is the proximal colon. After a short section of about 1 to 1.5
cm carrying the only striated muscles of the digestive tract and called
fusus coli, the wall becomes smooth in its terminal part; this part
is called the distal colon. Its last part is called the rectum
and ends at the anus. The latter carries the anal glands. |
| |
|
|
|
| |
|
|
The
digestive tract is relatively more developed in young rabbits than in
adults. It reaches its definitive size when the rabbit weighs 2.5-2.7
kg, a weight which is only 60-70 % of its adult weight. Two vital organs
release their secretions into the small intestine: the liver and
the pancreas. Hepatic bile contains salts and various organic substances,
but no enzymes. It indirectly aids digestion. In contrast, the pancreatic
juice contains a considerable amount of digestive enzymes which permit
protein (trypsin, chymotrypsin), starch (amylase) and fat (lipase) degradation.
In addition to their digestive functions, liver and pancreas have very
important functions in relation with rabbit's general metabolism |
| |
Conclusion
on the anatomy of the digestive tract |
| |
 |
|
|
Generally
speaking, the length of the small intestine (3-3.5 m) must be
underlined as well as its low relative content. The importance
of the role of the stomach and caecum as reservoirs must also
be underlined : 70 - 80 % of the total dry content of the digestive
tract is distributed between these 2 reservoirs. Finally, the
percentage of water in the digestive content may vary noticeably
from one segment to another as a result of secretions from the
organism, and water absorption.
|
|
| |
4.2
Digestive development according to age and physiological status.
|
| |
|
|
|
|
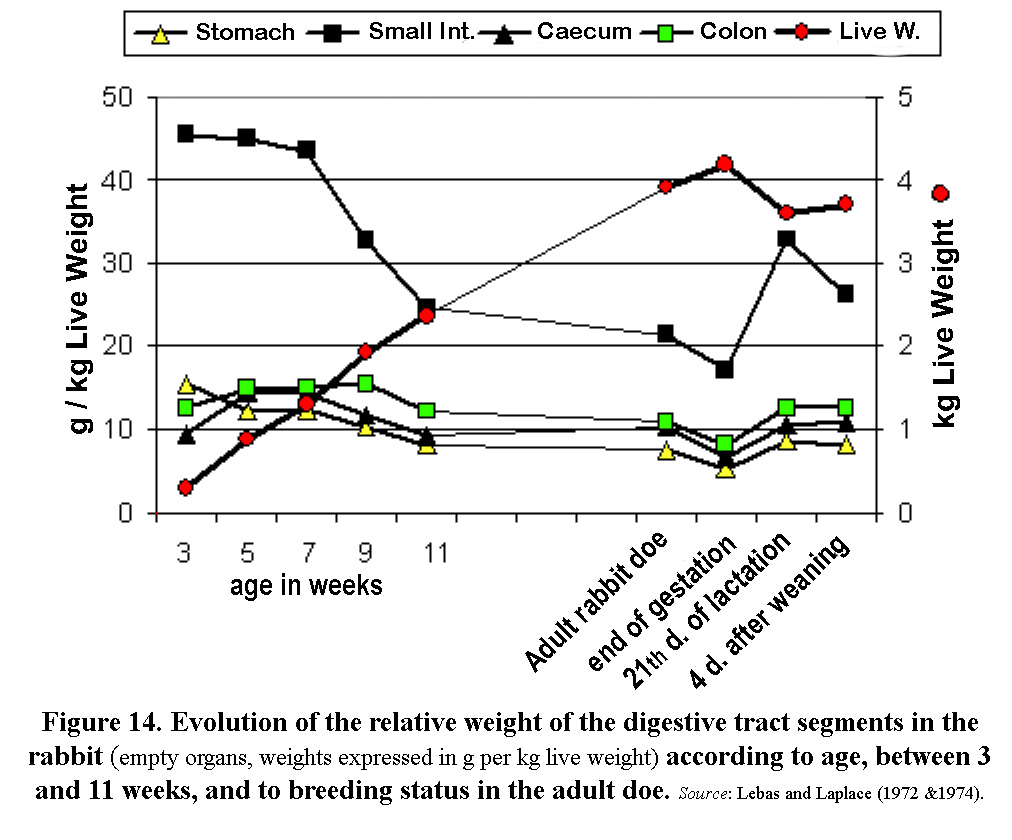
Between 3
and 5 weeks of age the anterior segments of the digestive tract
(stomach and small intestine) become less important, whilst the
posterior ones increase significantly (figure 14). From the age
of 7 weeks, the organs in the digestive tract develop less rapidly
in relation to the live weight of the rabbit, in particular the
colon which is always delayed by two weeks. In adult does the
importance of the digestive tract decreases at the end of gestation
but rapidly increases during lactation until it reaches a relative
size similar to that of a 5-7-week-old rabbit, (figure 14), an
evolution which demonstrates its plasticity.
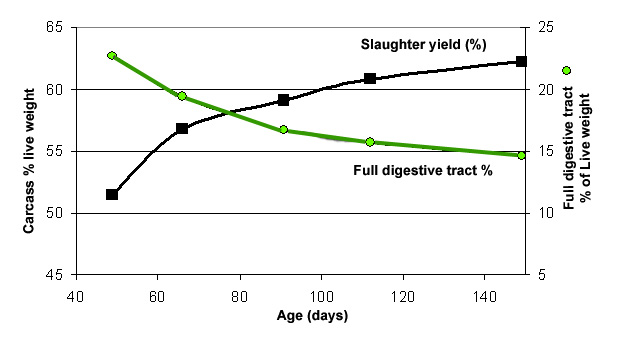
Taking into
account the evolution of the digestive mass and the relative proportionality
existing between the weight of the organ and that of its contents,
the slaughter yield systematically improves with age, at least
for kits older than 6 weeks (figures 15 and 16). The weight of
the raw skin, which is another important element of carcass yield,
only increases slightly from 14.5% up to 15.1% of the live weight
between 50 and 150 days, whereas the digestive mass decreases
in the sames interval of age from 23 % of the live weight at 50
days, to less than 15 % at 150 days (Combes et al., 2000). The
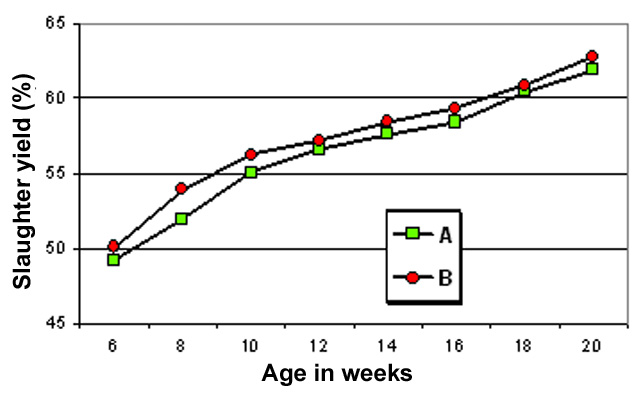
evolution of slaughter yield may be a little bit different from
one rabbit line to the other ((Figure 16)
|
| Figure
14. Evolution of the relative weight of the digestive tract segments
in the rabbit (empty
organs, weights expressed in g per kg live weight) according
to age between 3 and 11 weeks, and according to the breeding status
of the adult doe. Source: Lebas and Laplace (1972 and
1974). |
|
| |
|
|
|
|
|
|
Figure
15. Evolution of the relative weight of the full digestive tract
and slaughter yield, in a commercial strain of kits slaughtered
between 50 and 150 days (Source: Combes et al., 2000).
|
Figure
16. Evolution of slaughter yield with the age, in two experimental
strains A and B. Note:
mean of 30 kits was used for each age (an equal number of both
sexes) - mean weight of strain A at 6, 10 and 20 weeks: 1.355,
2.600 and 5.000 g ; mean weight of strain: B at the same ages:
1.455 -, 2.550 and 4.750 g (strain B is more immature). Source:
Lebas and Retailleau (1999)
|
|
| |
4.3
Physiology of digestion and caecotrophy |
| |
Digestive
transit |
| |
|
|
Consumed
feed particles swiftly reach the stomach where they find a very acid medium
and remain there for a few hours (about 2-4) without undergoing very significant
chemical changes. Marked acidification occurs, which causes various substances
to solubilise and beginns protein hydrolysis as a result of the action
of the pepsin. The stomach content is progressively «injected»
into the small intestine in the form of small waves caused by strong stomach
contractions. From the moment it enters the small intestine, the content
is diluted by the bile flow, the first intestinal secretions and, finally,
by the action of the pancreatic juices. Enzymes contained in these secretions
enable the release of easily absorbable elements which penetrate the intestinal
wall and are transported to the cells via the blood. After remaining in
the small intestine for about 1.5 h, the particles that have not been
degraded enter the caecum where they remain for some time (2-12 h), during
which they are attacked by the bacterial enzymes in the caecum. The elements
issued of the degradation by this new method of attack (mostly volatile
fatty acids) are released. They then penetrate the digestive tract wall
and are absorbed into the blood. |
| |
|
|
The content of the
cecum in turn is evacuated to the colon. Approximately half of the content
is coarse and fine feed particles that were not previously degraded.
The other half is bacterial bodies that have developed in the caecum
at the expense of the elements coming from the small intestine, as well
as the remains of digestive secretions also from the small intestine.
|
| |
The
alternative functioning of the proximal colon: basis of the duality of
excretion |
| |
|
|
Tiil this point
, the function of the digestive tract of the rabbit is no different
from other monogastrics. However, the dual nature of the functions
of the proximal colon is unique. If the content enter the colon
early in the morning, it undergoes few biochemical transformations.
The colic wall secretes a mucous which progressively envelopes the pellets
formed by the contractions of the large intestine wall. These pellets
form an elongated cluster, called soft faeces or "caecotrophs".
It is a different matter if the caecal content passes into the colon
at another time during the day. In this case, the colon contracts in
alternating directions; some of these contractions tend to empty the
contents "normally" and others send it back towards the caecum.
Due to the differences in the power and speed of movement of these contractions,
the content is squeezed out like a sponge. Most of the liquid fraction,
which contains soluble products and the fine particles (smaller than
0.1 mm, a dimension which includes the bacteria), is brought back to
the caecum, whilst the "solid" fraction containing the coarse
particles (larger than 0.3 mm), forms the hard faeces which are excreted
out of the rabbit (Björnhag, 1972). In fact, thanks to this dual
function the colon produces two different kinds of faeces: hard faeces
and caecotrophs. Their chemical composition can be seen in table 4 below
|
|
|
|
|
|
Caecotrophs
and hard faeces
(drawing from nature)
|
Twa
hard faeces and one lost caecotroph on the soil of a cage in a
raising on meadow system
|
Hard
faeces observed under the cage of a little bit perturbated rabbits,
some intermadiary soft faeces can bee seen,but no true caecotroph
obsrvable
|
Hard
faeces (classical droppings) of wild rabbits, observed in a field
|
|
| |
|
|
|
Table
4 : Chemical composition of the rabbit's faeces
Means and variablity
for 10 experimental diets, including complete diets and forages
(according to Proto, 1980)
|
| |
Hard
faecs
|
Soft
faeces (caecotrophs)
|
| |
mean
|
extremes
|
mean
|
extremes
|
| -
Dry matter (%) |
53.3
|
48-66
|
27.1
|
18-37
|
|
in
% of dry matter
|
|
|
|
|
| -
proteins |
13.1
|
9-25
|
29.5
|
21-17
|
| -
crude fibre |
37.8
|
22-54
|
22.0
|
14-33
|
| -
lipids |
2.0
|
1.3-5.3
|
2.4
|
1-4.6
|
| -
minerals (ash) |
8.0
|
3-14
|
10.8
|
6-16
|
|
| |
|
|
Difference between caecotropy and coprophagy clic
on the button 
|
| |
|
|
If the hard faeces
are evacuated in the litter, conversely, the caecotrophs are recovered
by the animal as soon as they are released from the anus. To this end,
during the emission, during an overall grooming operation, (Faure et
al, 1963) the rabbit turns around, sucks up the soft faeces as soon
as they come out of the anus, then swallows them without chewing them.
Therefore, the rabbit can, without any inconvenience, practice the recovery
of caecotrophs even if it is on a wire mesh floor. This is why if a
breeder observes cecotrophs under the cages of his rabbits, it means
that the animals are disturbed. In case of accumulated litter under
wire mesh cages, the hard droppings roll over each other when they reach
the ground and thus form "spread" piles. If caecotrophs are
not picked up by the animal due to a temporary or permanent stress,
the mucus around them tends to "stick" the droppings to each
other. In this case, the pile of droppings under the cages (directly
below the hindquarters of a rabbit when it is consuming in the feeder)
is then of "pointed" shape.
|
| |
|
|
When
everything is functioning normally, large quantities of soft faeces can
be found in the stomach, where they account for as much as three quarters
of the content (Gidenne and Lebas, 1988). The caecotrophs are digested
in exactly the same way as "normal" feed. Bearing in mind the
eventually recycled fractions, once, twice, even 3 or 4 times, and depending
on the type of feed, the digestive transit of the rabbit lasts about 15
to 30 hours (20 hours on average). The general functioning of the digestive
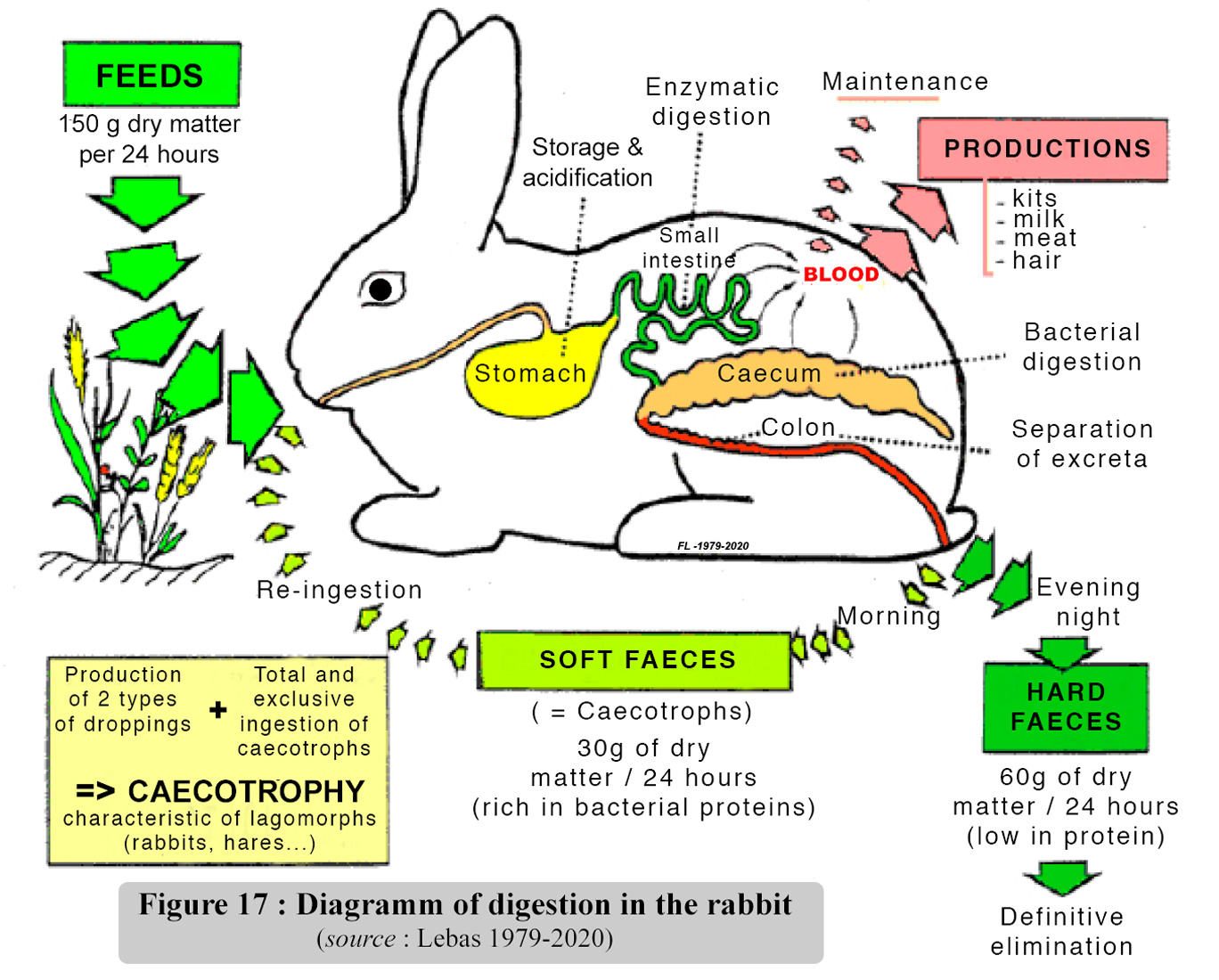
tract is summarized in figure 17. |
| |
|
|
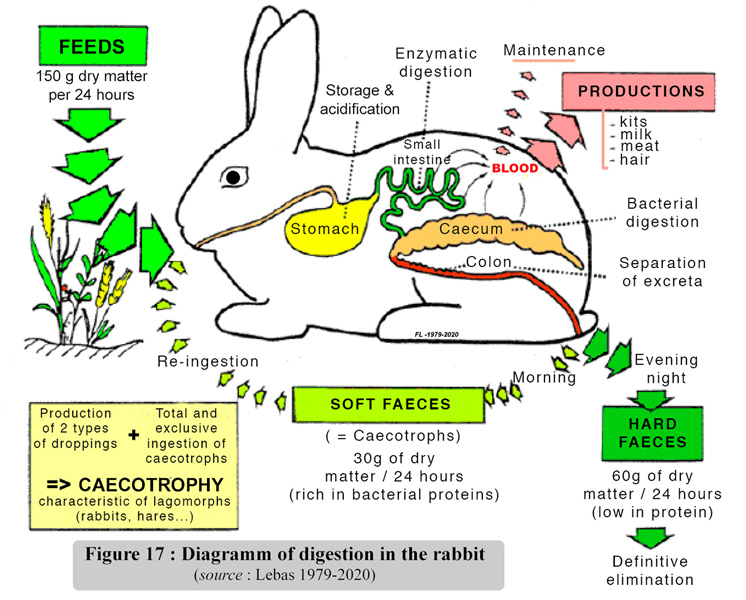 |
| |
|
|
It is worth remembering
that half of the caecotrophs content is made of bacterial bodies and
the rest is constuted by partially degraded feed residues and by the
remains of secretions from the digestive tract. The bacterial bodies
are an important source of proteins of high biological value, together
with hydro soluble vitamins. Caecotrophy practice, a priori, is of considerable
nutritional value. In a healthy rabbit fed on a balanced diet, caecotrophy
provides approximately 15 - 25 % of the daily protein intake and covers
the total requirements of vitamins B and C . However, this type of functioning
of the digestive tract and the quantities involved limit the quantitative
impact on protein nutrition, on the contrary the contribution of caecotrophy
to the hydro soluble vitamins supply is essential. The administration
of vitamins is often advised, for example during the days following
weaning, because of the possible risk of digestive disorders which stop
the hydrodulublrd vitamins supply.
|
| |
|
|
|
This
peculiar way of functioning of the digestive tract requires a
minimum content of ballast such as crude fibre. In fact, if the
feed contains few coarse particles, or is highly digestible, or
both, transit to the caecum is very fast but the caecal content
stay a long time in this organ, and becomes poorer in elements
that may feed the " normal " bacteria living in this
part of the digestive tract. So, it seems evident that different
other bacteria can proliferate in this poor medium and frequently
some of them may be harmful. It is therefore necessary to provide
a minimum of ballast in the feed for reduced caecal retention
time and promote a rapid transit. Dietary ballast has been in
the past, mistakenly associated only with crude fibre in feeds.
At present, minimum amounts of ligno-cellulose (ADF) and lignin
are recommended, because they better discribed the "ballast"
effect of the feeds.
|
|
| |
|
|
Figure
18 shows the influence of the amount of fibre ingested (fixed composition),
on the feed retention time in the different segments of the digestive
tract. It should be remembered that the effects on retention times in
the stomach and small intestine tend to compensate each other but the
differences between extreme feeds do not exceed 2 hours. On the other
hand, a reduced supply of fibre has more effect on the time during which
the bolus is retained in the caecum: between 9h 40 min. and 21 h 30 min
if the lowest fibres quantity is taken in consideration |
| |
|
|
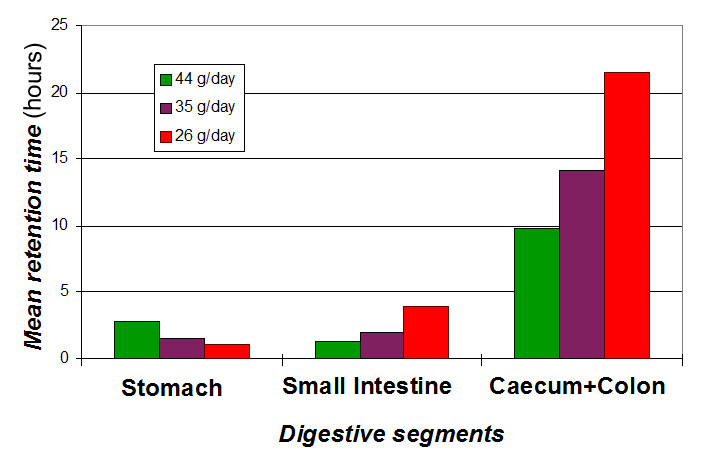 |
| Figure
18. Retention time (hours) in the different digestive segments
(stomach, small intestine and caecum + colon), after the ingestion
of controlled amounts of fibre (NDF): between 26 and 44 g/day.
Source: Gidenne (1994). |
|
| |
|
|
Caecotrophy regulation
depends on the integrity of the digestive microbiota as well as the
ingestion rate. The ingestion of caecotrophs starts 8 - 12 hours after
feeding in restricted animals or after the ingestion peak in animals
fed ad libitum. The ingestion rate in animals fed ad libitum
and consequently the caecotrophy rate, are the result of the light period
to which they are subjected (see
below Feeding behaviour). It should also be pointed out that
caecotrophy depends on internal regulation mechanisms which are not
completely known today . For example, adrenal gland ablation implies
that the animal no longer practises caecotrophy; the administration
of cortisone to adrenalectomized animals enables normal caecotrophy
behaviour to be resumed. It therefore appears that digestive transit
in rabbits is dependent on adrenalin secretions. Stress-related hypersecretion
causes hypoperistalsis (which
increased retention time , effect similar to the effect of a low fibre
diet) and as a consequence promote a high risk of digestive disorders
.
|
| |
|
|
Caecotrophy
is observed in young kits (domestic or wild) around 3 weeks old, as soon
as they start consuming solids in addition to the mother's milk. |
| |
|
|
|
| |
4.4
Feeding behaviour
|
| |
|
|
Feeding
behaviour has been studied mostly in rabbits fed a complete pelleted diets,
or in studies on feed preferences using dry or fresh feeds (grains, straws,
hay, roots, amongst others). However, various studies of rabbits in semi-freedom
and wild rabbits provide a better understanding of the feeding behavior
of domestic rabbits raised conventionally in cages or in parks.
|
| |
Ingestion
rate |
| |
|
|
Suckling
rabbits |
| |
|
|
The first feeding
of a young rabbit is very usually done during parturition itself. After
parturition, the doe herself decides the suckling rate for her kits
and she feeds them once a day (Cross, 1952). Suckling lasts 2-3 minutes
and the female does not provide direct assistance to the young, she
is content to position herself correctly above the litter to give good
access to all the teats. Some does occasionally feed twice a day or
visit the nest several times, leading "observers" to believe
that lactation occurs 4-5 times a day. Multiple suckling is not interesting,
as already demonstrated by Zarrow et al. (1965), when growth rate of
kits fed by the mother once or twice a day were found to be identical
to that of the kits fed during unrestricted visits to the nest.. These
results were confirmed more recently for example by Tudela and Balmisse
in 2003 who showed that if 2 feedings per day allow the young rabbits
to obtain a greater volume of milk (+ 8%), but the total quantity of
nutrients obtained is the same since the average weight of the young
rabbits at 21 days is strictly identical in the 2 situations as had
been previously demonstrated (Table 5).
|
| |
|
|
| Sucklings
per 24 h |
one
|
two
|
| Number
of observed litters (9 kits /litter) |
70
|
70
|
| Milk
production in 24h ( mean of days 2 to 21) |
250
g/day
|
271
g/day
|
| Average
weight of one kit at 21 days |
394.3
g
|
393.9
g
|
| Table
5 : Milk production of rabbit does authorized to
feed their litter of 9 kits, once or twice per 24 h.
Two
consecutive litters observed per doe, milk suckling rhythm was
alternated between the 2 successives litters. Source: Tudela
and Balmisse (2003). |
|
| |
|
|
Sometimes,
if the amount of milk is insufficient, the kits try to suckle every time
the mother enters the nest, but she retains the milk. This behaviour is
characteristic when milk production is insufficient. |
| |
|
|
Conversely, if the
rabbits are offered to suckle twice a day at 12-hour intervals, but
with a different mother, one in the morning and another in the evening,
they readily accept. They can then ingest more milk (+ 37% on average,
but ingestion is not x 2) and benefit from it for their growth (Table
6). It is therefore effectively the mother who determines the rhythm
and the quantity of milk available to the young rabbits.
|
| |
|
|
| Number
of rabbit does nursing the same litter |
One
|
Two
|
| Test
1 - number of litters (equalized at 8 at birth) |
50
|
25
|
| Milk
/ kit/ 24 hours (weeks 1-3) |
24.6
g/d.
|
33.6
g/d.
|
|
Average
weight of 1 kit at 21 days
|
300
g
|
466
g
|
| Test
2 - number of litters (equalized at 8 at birth) |
33
|
27
|
| Milk
/ kit/ 24 hours (weeks 1-3) |
25.6
g/d.
|
35.7
g/d.
|
| Average
weight of 1 kit at 21 days |
342
g
|
446
g
|
Table 6: Average
milk consumption and weight at 21 days of young rabbits sucking either
only their own mother in the morning,
or sucking their own mother in the morning and a second rabbit doe
12 hours later. Source:
Szendrö et al. (2000)
|
| |
|
|
During
the third week of life, the kits begin to move in a perfectly coordinated
manner. They ingest milk, a little drinking water, + a very small quantity
of mother's feed if available, + some hard faeces deposed by the mother
in the nest at their intention in order to provide an adult rabbit's mictobiota
to her kits. During the fourth week, kits ingest more solid feed and water
than milk. Changes in feeding behaviour at this period are truly extraordinary:
the young rabbit under the mother passes from a single feeding per day
to a multitude of solid and liquid meals more or less alternated and distributed
irregularly throughout the day, rythm characteristic of the feeding behavior
of the adult. It is also during of this 4th week of life that begins the
practice of cecotrophy. In fact, in the stomachs of young rabbits sacrificed
at 22 days, only milk and feed are found; while in young rabbits sacrificed
at 28 days cecotrophs can be identified in the stomach in addition to
feed and traces of milk (for details see the article of Orengo
& Gidenne, 2005). The preferentially nocturnal ingestion of the
solid feed is already marked. |
| |
|
|
|
| |
|
|
|
| |
|
|
It
is interesting to note that when it begins to consume solid feed, the
suckling rabbit has a clear preference for maternal feed even over feed
which is better suited to its physiological needs. This suggests a role
for the mother in learning to eat feed, but this has not been formally
demonstrated. However, by playing on the appropriate flavoring of the
"young rabbit" feed, it is possible to encourage them to consume
more of it than maternal feed (table 7). |
| |
|
|
| The
2 free choice feeds => |
Maternal
feed |
Flavoured
"young" feed |
| Aroma
test N°1 (21 to 35 days) |
|
|
| Average
intake / kit |
40.4
g/day |
7.9
g/day |
| Relative
intake |
83% |
17% |
| Aroma
test N°2 (21 to 30 days) |
|
|
| Average
intake / kit |
8.1
g/day |
13.1
g/day |
| Relative
intake |
38% |
62% |
Table 7: Attempt
to flavor the feed intended for young rabbits before weaning :
feed intake by the young rabbits in a situation of free choice,
the mother not having
access to any of the 2 feeds. Source: Mousset, 2003.
|
| |
|
|
Feeding
behaviour of he weaned rabbit or of adult rabbit |
| |
|
|
At
the time of weaning, the young rabbits already have 30 to 40 solid or
liquid meals per 24 hours (table 8). The total time spent on meals in
a 24 hour cycle is, at 6 weeks, more than 3 hours. Then rapidly it drops
to under 2 hours. If the rabbit is offered a non-pelleted feed (flour
or mash), the time spent eating is doubled. Whatever the animals' age,
feed containing over 70 % water (green roughage or roots, for example)
at a temperature of 20 ºC, provides more than the required amount
of water. |
| |
|
|
| |
Age
in weeks
|
| |
6
|
9
|
12
|
15
|
18
|
|
Live
weight and growth rate
|
| -
weight of rabbits (g) |
1060
|
2094
|
2922
|
3532
|
3901
|
| -
ADG g/day |
49.2
|
44.3
|
34.3
|
23.3
|
17.6
|
|
Solid
feed intake (pellets , 89% DM)
|
| -
g pellets / 24 h |
98
|
168
|
194
|
184
|
159
|
| -
N° of meals / 24h |
39
|
39
|
41
|
41
|
34
|
| -
g / meal |
2.6
|
4.4
|
4.9
|
4.4
|
4.7
|
|
Drinking
water
|
| -
g (ml) water / 24h |
153
|
269
|
320
|
319
|
298
|
| -
N° of drinks / 24h |
31
|
26
|
29
|
31
|
36
|
| -
g (ml) water / drink |
5.1
|
10.4 |
11.5
|
10.8
|
9.1
|
|
Water / Feed ratio |
1.56
|
1.60
|
1.65
|
1.73
|
1.87
|
Table 8. Evolution
of the feeding behavior of male rabbits between 6 and 18 weeks,
having permanently at their disposal a complete pelleted feed and
drinking water, kept in a room at 20 ± 1°C. Source:
Prud'hon et al. (1975).
|
| |
|
|
The
distribution of feed and water intakes is not homogenous during the 24
hours period (figure 19). The proportion of daily feed consumed on an
hourly basis during the darkness period is greater than that ingested
during the light period, for both solids and liquids. It should be pointed
out that consumption is higher just before the lights are switched
off in the raising room. In sub adult rabbits (3 kg New Zealand White)
with 12 hours of light/24 h., nocturnal consumption represents almost
two thirds of that observed in the total 24 hours cycle, due to increased
feeding frequency but the amount of each meal is the same, 5-6 g per meal.
|
| |
|
|
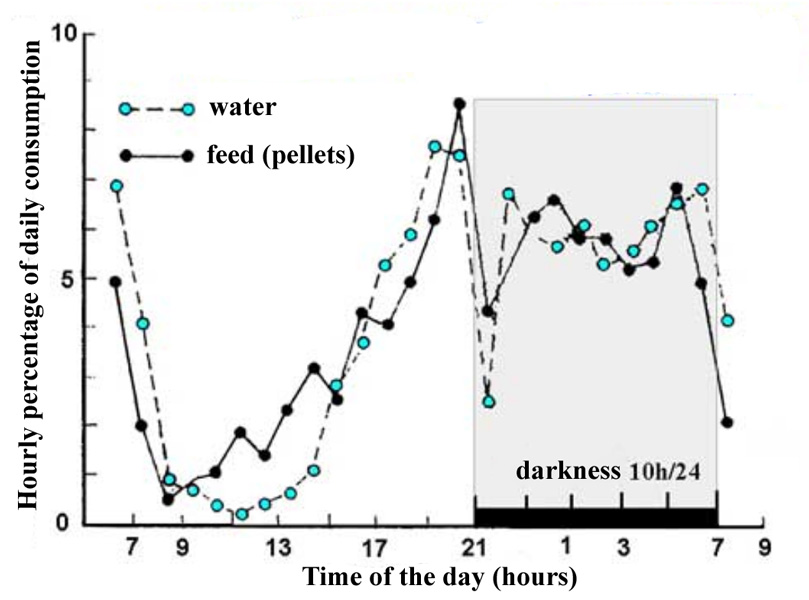
Figure 19. Hourly distribution of wáter and pelleted feed
consumption over a 24-hour cycle in a 12 week old rabbit.
Source: Prud'hon et
al (1975). |
| |
|
|
As
the rabbits grow, their nocturnal feeding behaviour increases. Feed intake
decreases during the light period and the morning,"feeding rest"
period between meals becomes longer (figure 20). The feeding behaviour
of wild rabbits is even more nocturnal than that of domestic ones |
| |
|
|
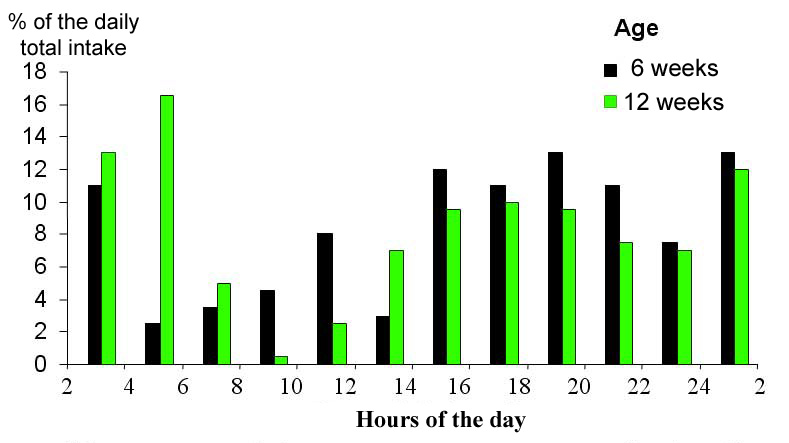
Figure 20. Distribution of daily feed consumption in 2 hours increments,
in 6 and 16 week old rabbits.
Average consumption of 80 and 189 g of pelleted feed per day for the
2 ages. Lighting 12 hours a day from 7 a.m. to 7 p.m.
Source: Bellier et
al. (1995). |
| |
Evolution
of feed ingestion in terms of age and physiological status |
| |
|
|
The
amount of feed and water ingested depends firstly on the nature of the
feed given to the rabbits at a given time, and especially on the digestible
protein and energy content: high energy content tends to reduce consumption
and high protein content tends to increase it . These amounts also depend
on the type of animal, its age and breeding status or on the ambient temperature.
|
| |
|
|
Consumption
of kits depends to a large extent on the animal's age, as well as other
factors (figure 21). Using spontaneous consumption in adults as reference,
(for example, 140-150 g DM/day, in New Zealand White rabbits weighing
4 kg), it was recorded that the daily consumption of a 4-week-old rabbit
is one quarter, whilst its live weight corresponds to only 14 % of the
adult's live weight. At 8 weeks the equivalent proportions are 62 and
42 % and at 16 weeks, 100 to 110 % and 87 %. |
| |
|
|
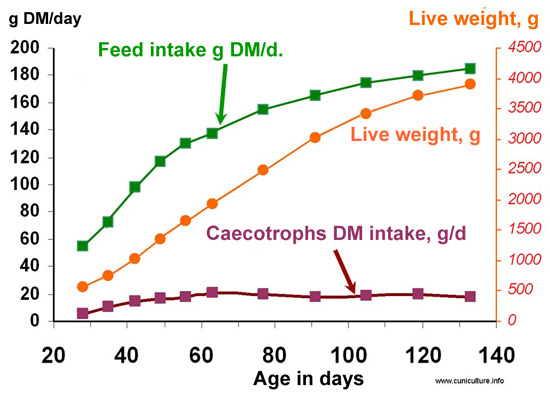
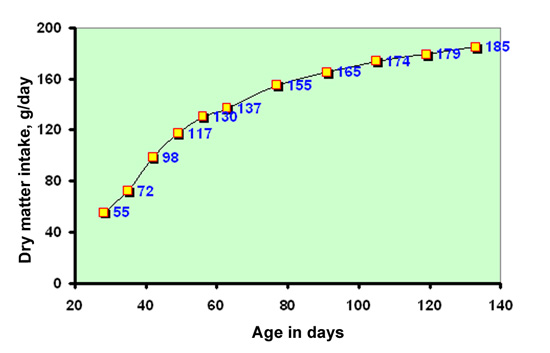 |
 |
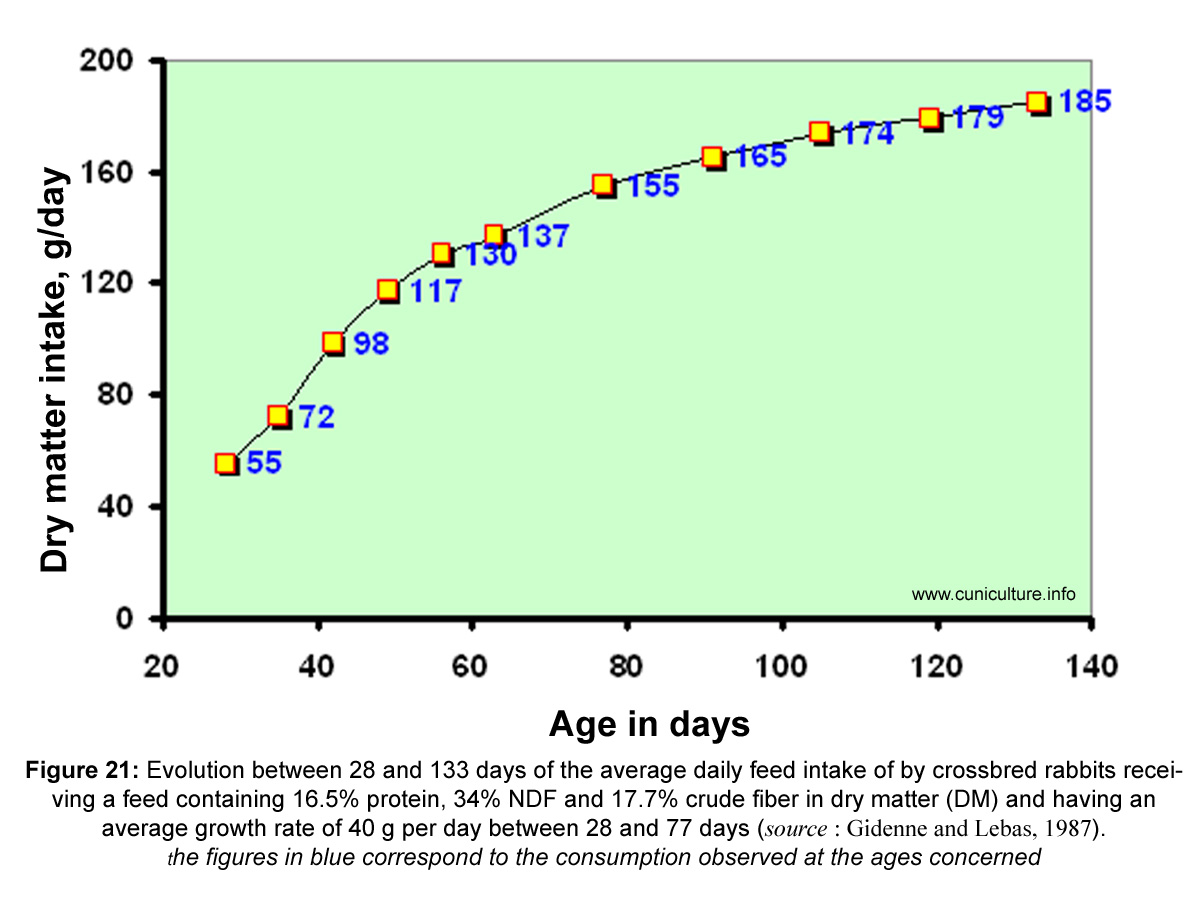
Figure
21: Evolution between 28 and 133 days of the average daily feed
intake of by crossbred rabbits receiving a feed containing
16.5% protein, 34% NDF and 17.7% crude fiber in dry matter (DM)
and having an average growth rate of 40 g per day between 28 and
77 days (source
: Gidenne and Lebas, 1987).
the figures in blue correspond
to the consumption observed at the ages concerned |
Figure
22: Comparative evolution of live weight and of feed (pellets)
and caecotrophs consumption between weaning and the adult stage
(Source Gidenne and Lebas, 2005) |
|
| |
|
|
The
rabbit regulates its ingestion according to its energy needs, like other
mammals. Chemostatic mechanisms are involved, through the nervous system
and blood metabolites linked to energy metabolism. However, in monogastric
animals glycemia plays a key role in the regulation of feed intake, while
in ruminants plasma concentration in volatile fatty acids plays an important
role. Given that the rabbit is an herbivorous monogastric, glycemia seems
to play a preponderant role in relation to the concentration of VFA, but
the respective role of these two metabolites (glucose vs VFA) on the regulation
of ingestion remains today poorly understood. |
| |
|
|
Voluntary
ingestion is in fact proportional to metabolic body weight (PV0.75,
and is approximately 900-1000 kJ ED / day / kg PV0.75 (ED:
digestible energy). Chemostatic regulation would intervene beyond a DE
concentration of 9 to 9.5 MJ / kg. Below this level, a physical type regulation
prevails which would be linked to the state of fullness of the digestive
tract. |
| |
|
|
The
ingestion of caecotrophs increases up to 2 months of age and then remains
stable (figure 22). Expressed in fresh matter, it evolves from 10 g/ d
to 55 g/ day between 1 and 2 months of age, and represents 15 to 35% of
feed intake. However, it is possible that these values are underestimated
given the measurement technique used in this work (temporary installation
of mini shackles preventing re-ingestion of caecotrophs). |
| |
|
|
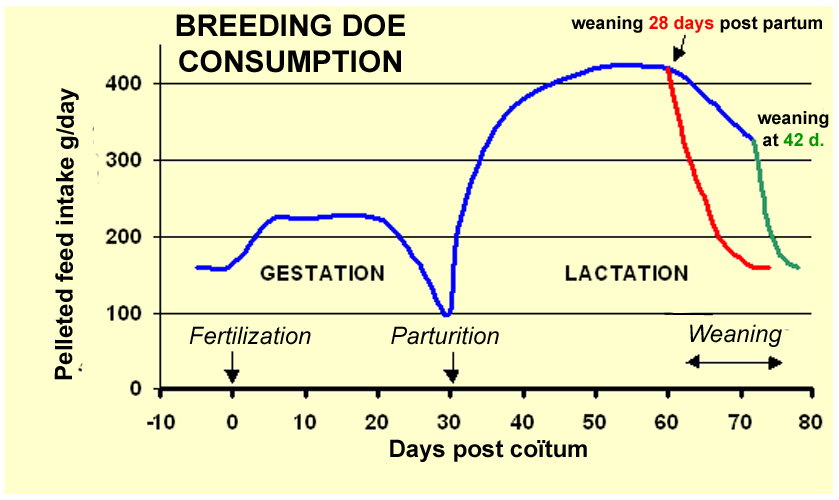
Spontaneous
feed consumption in rabbit females varies during the reproduction cycle
(figure 23). Decreased consumption at the end of gestation is marked in
all does and in some cases, solid feed intake can stop altogether on the
eve of parturition. In contrast, water intake never stops. After parturition,
feed ingestion increases rapidly and can exceed 100 g of dry matter per
kg of live weight. Water intake is also significantly increased at this
time: 200 to 250 g/day per kg of live weight, i.e. up to one liter
/day for a 4 kg rabbit doe. When a doe is simultaneouly gestating and
lactating, her feed consumption is comparable to that of a only lactating
doe, and never exceeds this amount. |
| |
|
|
|
|
|
Figure
23. Evolution of feed intake in the rabbit doe. Pelleted dry
feed, with 89 % dry matter.
Study carried out during gestation and subsequent lactation.
Source: Lebas (1975).
|
|
| |
Ingestion
of feed and water depending on the environment |
| |
|
|
Effect
of temperature |
| |
|
|
The
amount of energy a rabbit uses up, depends on environmental temperatures.
Feed intake compensates for this and therefore also depends on the environmental
temperature. Laboratory work carried out shows that when temperatures
increases from 5ºC up to 30º, the consumption of growing rabbits
is reduced from 180 g down to 120 g/day for dry feed, and water ingestion
is increased from 330 up to 390 g/day in the same conditions (table 9).
A more precise analysis of behaviour shows
that when the temperature increases, the number of meals / 24h. (solid
and liquid) decreases . For example young New Zealand females go from
37 solid meals at 10º C down to 27 meals /day at 30º C. On the
other hand, if the amount of feed consumed in each meal decreases as a
result of high temperatures (5.7 g/meal at 10° C and 20° C compared
to 4.4 g at 30° C), the opposite occurs for water , the amount consumed
each time increases with the temperature increase (from 11.4 up to 16.2
g per meal, at 10 °C and 30 °C). |
| |
|
|
| Temperature |
5°C
|
18°C
|
30°C
|
| Relative
Humidity |
80%
|
70%
|
60%
|
| Pelleted
feed intake |
182
|
158
|
123
|
| Water
intake |
328
|
271
|
386
|
|
Water/Feed ratio |
1.80
|
1.37
|
3.14
|
| Daily
weight gain |
35.1
|
37.4
|
25.4
|
Table
9. Feeding behavior of growing rabbits as a function of the ambient
temperature.
Consumption and weight gain in g/ day.
Source: Eberhart (1980).
|
| |
|
|
If
the consumption of growing rabbits is affected at 30 °C and above
, that of breeding rabbits is equally affected as shown in figure 24.
It should be noted that milk production is also affected by heat in the
same proportion as the consumption of pelleted feed (at 30°C milk
production is 70% of the value measured at 23° C).
|
| |
|
|
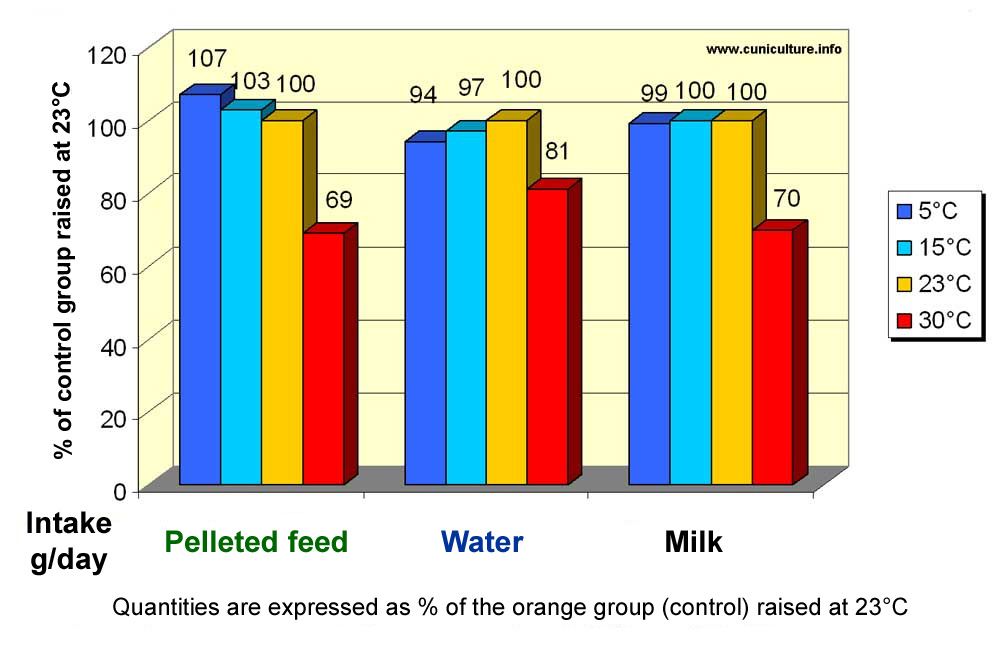
Figure 24: Effect of ambient temperature on feed and water intakes,
and on rabbit milk production.
(Source: Szendrö
et al., 1998) |
| |
|
|
A
study carried out in Italy by Finzi et al. (1992), shows that if the temperature
increases (tests carried out at 20 °C, 26 °C and 32 °C), the
ingested feed/water ratio increases significantly, which is nothing new.
However, the different ratios between ingestion and excretion also change
(table 10). These authors propose using these ratios (the ones easiest
to calculate locally), to verify the existence of thermal stress in rabbits.
However, this suggestion should be validated before implementation. |
| |
|
|
| Ratios |
Temperature
|
|
20°C
|
26°C
|
32°C
|
|
Water
/ Feed
|
1.7
|
3.5
|
8.3
|
|
Urine
/ Feed
|
1.0
|
1.6
|
1.0
|
|
Water
/ Faeces
|
1.9
|
5.5
|
11.2
|
|
Urine
/ Faeces
|
1.1
|
2.5
|
5.3
|
Table 10. Incidence
of environmental temperatures on different traits relative to
ingestion and excretion in adult rabbits.
Source: Finzi et al. (1992).
|
| |
|
|
Relation
Water-Feed |
| |
|
|
|
If a rabbit
has no access to drink water at all and there is only dry feed
available (less than 14 % water), dry feed intake will stop completely
within 24 hours. Under these conditions of water lack and depending
on the environmental temperature (temperature and hygrometry),
an adult rabbit can survive 4 to 8 days without its vital functions
undergoing any irreversible alterations. However its weight can
decrease by 20 to 30 % in less than a week. If only clean water
is freely available a rabbit will survive 3 to 4 weeks without
solid feed. As for what is considered as "normal", water
ingestion increases 4 to 6 fold in a few days. Sodium chloride
distributed in the water (0.45 %) reduces this increase in consumption;
but potassium chloride does not (sodium is lost through the urine,
not potassium). Rabbits are therefore very resistant to fasting
and relatively resistant to lack of water but it should be pointed
out that any limitation to the amount of water in relation to
the rabbit's needs, means a proportional drop in the dry material
ingested, and consequently, alterations in productivity.
In some cases,
decreased water ingestion occurs because the animals cannot reach
the drinking troughs. Suckling kits sometimes have to climb onto
the mother to do this because they are so high up (over 25 cm
between the floor of the cage and the waterer). As long as they
ingest milk (70 to 75 % water in the milk) and the mother is tolerant,
the kits will have no problems with the drinkers. But the situation
can become dramatic after weaning, (no more mother to climb on)
especially in the case of early weaning (before the end of the
4th week). The risk is further magnified for small breeds.
|

Figure
25: The dinker must be accessible for the young rabbits: 25
cm maximum from the floor of the cage and close to a wall
|
|
| |
|
|
Effect
of other environnmental factors |
| |
|
|
Other
environmental factors have also been studied in domestic rabbits, such
as the lighting schedule or housing systems. In the absence
of light (24 hour darkness), the ingestion of the growing rabbit is slightly
increased in comparison with rabbits subjected to a light program with
a 24 hour cycle. In the absence of light, the rabbit organizes its feeding
program on a regular cycle of 23.5 to 23.8 hours, with 5 to 6 hours devoted
to the ingestion of caecotrophs. In continuous lighting, the feeding program
of the rabbit is organized on a cycle of approximately 25 hours, and the
total feed intake / 24h is reduced and the growth performance too. |
| |
|
|
According to Hungarian
studies published in 2000, in the reproductive female, a modification
of the lighting program, by introducing 2 periods of darkness of 4 hours,
on cycles of 12h (light, darkness, 2 cycles/24h) reduces the ingestion
and causes an increase in milk production, thus leading to better feed
efficiency.
|
| |
|
|
As
mentioned earlier, the type of cage also influences the feeding behavior
of the rabbit. Ingestion is reduced if the density of rabbits in the cage
rises, possibly due to greater competition between animals for access
to the feeder, but also mainly due to reduced mobility of animals and
therefore of their nutritional needs (table 11). This density effect is
also observed for rabbits reared in individual cages (effect of cage dimension) |
| |
|
|
| Number
of rabbits / cage |
6
|
7
|
8
|
9
|
10
|
| Density
: rabbits / m² of cage |
16.9
|
19.6
|
22.6
|
25.4
|
26.2
|
| - initial
weight at 32 d. (g) |
773
|
773
|
772
|
770
|
772
|
| - average
daily gain 32-68 d. |
43.6
|
44.1
|
42.9
|
42.1
|
40.3
|
- consumption
32-42 d. (g)
|
100
|
99
|
97
|
98
|
99
|
| - consumption
42-55 d. (g) |
133
|
131
|
130
|
124
|
123
|
| - Consumption
55-68 d. (g) |
155
|
153
|
152
|
150
|
139
|
|
- Average
consumption 32-38d.(g/d)
|
132
|
130
|
129
|
126
|
122
|
| - Final
weight (kg) |
2.34
|
2.36
|
2.32
|
2.26
|
2.22
|
| - Final
density : kg liveweight / m² |
|
46.6
|
52.4
|
58.1
|
62.8
|
Table 11.
Feed consumption
and growth of rabbits between 32 and 68 days of age, at a density
of 6 to 10 subjects per cage of 0.354 m².
Source: Aubret and Duperray (1993); 8 cages per group.
|
| |
|
|
|
| |
|
|
| Rabbits/cage |
4
|
30
|
| 29-43d
(g/d) |
94
|
88
|
| 43-57d
(g/d) |
146
|
135
|
|
57-71d
(g/d)
|
164
|
164
|
| 29-71d
(g/d) |
135
|
125
|
Table
12. Consumption of rabbits raised in groups of 4 or 30, from 29
to 71 days of age, at the same density of 15.6 rabbits /
m².
Source: Maertens and Van Herck (2000).
|
Nevertheless,
rearing growing rabbits in very large collective cages (30 subjects
in 1.9 m²) authorize more movement for the animals which
reduce their daily ingestion compared to that of rabbits reared
at a rate of 4 per cage at the same animal density (125 vs 135
g / day). Unlike the effect of the number in the same cage (table
11), the reduction in consumption with large numbers of rabbits
in larges cagess at fixed density (15.6 rabbits / m²) is
evident even from the start of fattening (table 12).
Finally, the
number of places in front of the feeder (1 to 6 places) for a
group of 10 rabbits fed ad libitum, does not have a measurable
influence on the level of the pellets consumption. It is the same
for rabbits rationed not too drastically (at least 70% of consumption
at will).
|
|
| |
Feed
restriction, feeding behaviour and digestive development |
| |
|
|
It
may be necessary to limit the amount of feed given to growing rabbits
for several reasons. This is for example recommended in the event of epizootic
rabbitenteropathy, the ERE named EEL in French (see for example the articles
in French which were devoted to this topic in the magazine part of this
Website in 2003
or in 2009
or the report of the round table devoted to it by the Association Scientifique
Française de Cuniculture in 2007). |
| |
|
|
|
| |
|
|
Quantitative
rerstriction |
| |
|
|
When
a limited quantity of food is distributed to rabbits, the daily intake
is consumed all the more quickly as the restriction is more marked. For
example for rabbits housed in individual cages or in pairs, an allowance
representing 85% of the pellets consumed ad libitum, is completely
consumed in about 16 hours. If the allowance represents only 70% of the
consumption ad libitum, it is completely consumed in just under
10 hours. |
| |
|
|
In
the case of rabbits conventionally reared at a rate of 8 per cage, a pellets
allocation of 85% is completely consumed in 8 hours (98% in 5 hours -
figure 26) if only one rabbit can eat at a time (effect of competition)
while if two rabbits can eat simultaneously, only 89% of the same allowance
are eaten in 8 hours. For more details in French on the feeding behavior
of quantitatively rationed rabbits, see the article Tudela
and Lebas (2006) in the Magazine section of this Website. |
| |
|
|
|
|
|
Figure
25. Evolution of the cumulative intake during the 8 hours following
a single distribution in the morning at 8h00, of rabbits fed
ad libitum (control) or of rabbits receiving only 85%
of the ration of control rabbits in a feeder having only one
or two consumption stations for the 8 rabbits in the cage.
Source:
Tudela and Lebas (2006).
|
|
| |
|
|
Regardless
of the level of restriction, rabbits do not increase their instantaneous
rate of ingestion. They increase the duration of each meal, in particular
the one following the distribution, and reduce the interval between each
meal. The duration of these meals is however limited by the stomach capacity
which represents at most only about fifteen grams of pelleted feed for
a 2 kg rabbit, knowing that the stomach of a rabbit is never empty before
the start of a meal. This situation allows 8 rabbits to eat in turn, even
in a feeder with only one eating station. For example, even a 60% restriction
does not increase the variability in weight between the rabbits in a collective
cage, which means that each of the 8 rabbits was indeed restricted at
the same level and that none of them consumed the "neighbor's"
share. |
| |
|
|
|
| |
|
|
Restriction
of access time to the feeder |
| |
|
|
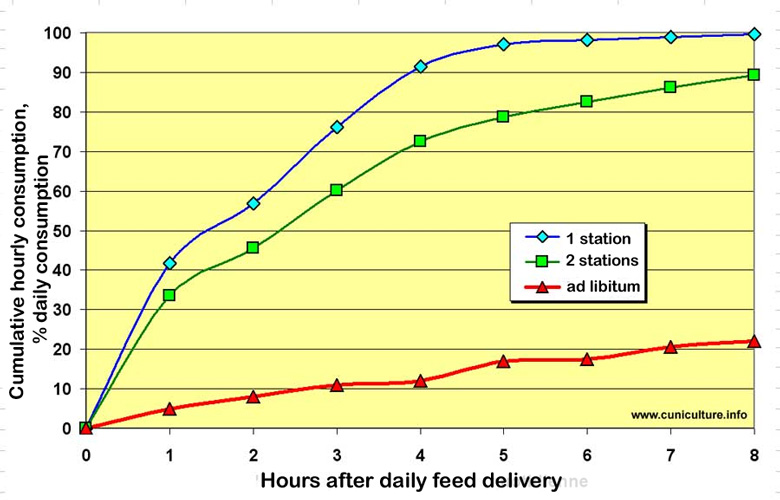
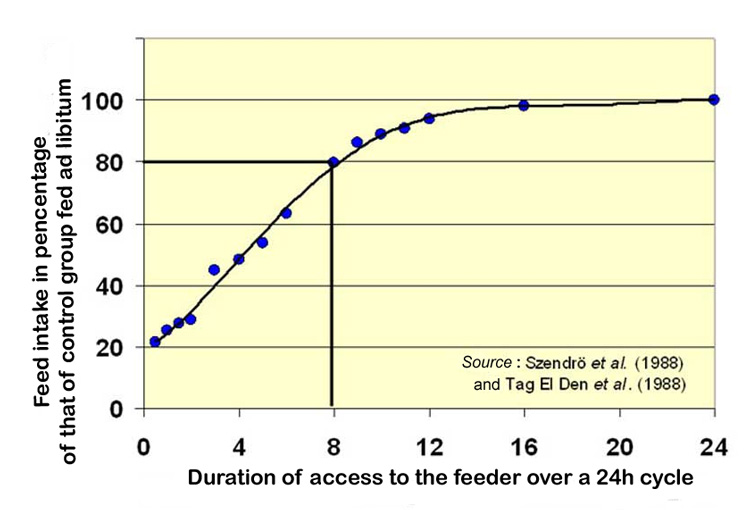
At
the end of the 1980s, the Hungarian team from Kaposvar under the direction
of Zs. Szendrö carried out a systematic study of the amount of feed
ingested by growing rabbits as a function of the time allowed for consumption
over 24 hours (a duration per 24 hour cycle varying from ½ hour
to 16 hours) in comparison with ad libitum fed rabbits (figure
27). So when rabbits have 8 hours a day to consume their ration, they
consume about 80% of what rabbits fed ad libitum consume. More
drastic reductions in access time result in an almost linear reduction
in the quantity of feed consumed, which is reduced to 20% for an access
time of only ½ hour/24h. |
| |
|
|
|
|
|
Figure
27. Feed consumption in 24 hours as a function of the duration
of access to the feeder, expressed as a percentage of the control
group fed ad libitum.
Source of the graph: ASFC 2007
|
|
| |
|
|
|
All these
results correspond to the average calculated over the entire growing
period after weaning. If, on the other hand, the evolution of
the effect of the restriction of the access time on the actual
consumption is of interest , it was seen that there is an adaptation
over time. For example, after 8 weeks of rationning the rabbits
arrive, even with only 8 hours of acces, to consume as much as
those fed ad libitum. This adaptation over time has been confirmed
by more recent works. For example, Foubert et al. (2007) have
shown that with an access time limited to 8 hours per day, the
effective restriction is 64% then 73% and finally only 83% for
each of the 3 consecutive weeks following a weaning at 32 days.
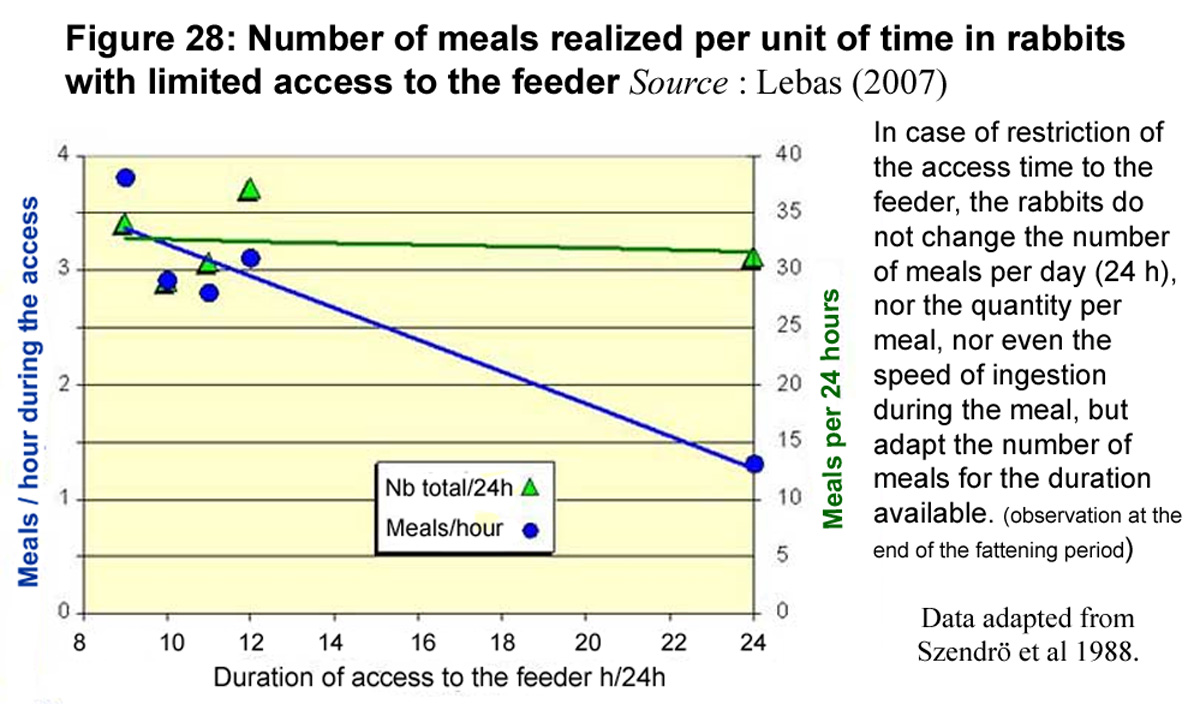
The Hungarian
team from Kaposvar also measured the number of meals made by the
rabbits according to the number of hours of access to the feeder
each day. It appears that, if they have at least 9 hours to consume
their solid feed, rabbits systematically realize 30 to 35 meals
per day. If they have all the day to eat, they eat at an average
rate of 1.3 meals per hour (figure 28). But if the access time
is restricted, this number of meals can increase to almost 4 meals
per hour, during "working" hours of course. On the other
hand, the duration of each meal is not significantly affected
by the duration of access to the feeder over 24 hours: 3.2 minutes
on average, nad the interval between meals is reduced. In total
over the day, 12-week-old rabbits fed ad libitum spend
about 1 hour 45 minutes eating, while if they have only 9 or 10
hours to consume their solid feed, they spend only 1 hour 20 minutes
approximately.
|
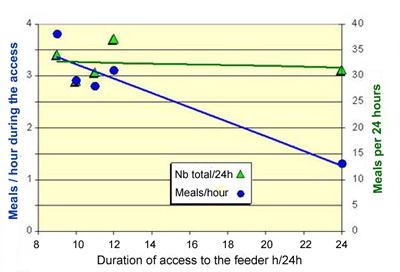
Figure
28 : Number of meals realized per unit of time in rabbits
with limited access to the feeder Source :
Lebas (2007)
|
|
| |
|
|
Restriction
of access time to the waterer |
| |
|
|
|
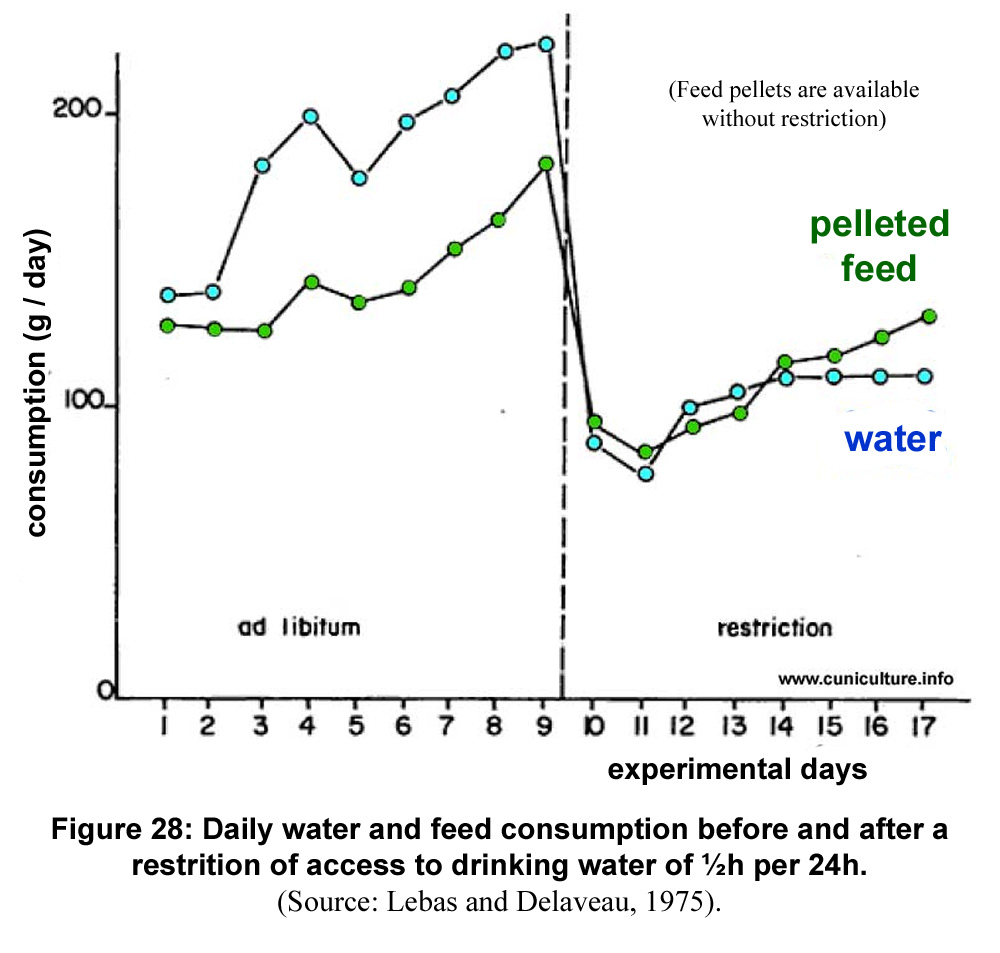
Work carried
a lot of years ago out at INRA in France at Montpellier (Prud´hon
et al., 1975) has shown that after one week of adaptation, 6-9
week old rabbits can access the waterer for only 10 minutes
each day. The 11-14 week old or adult rabbits respectively had
a pelleted feed consumption reduced to 86% of that of the control
group able to drink at will. Water consumption was reduced to
84% and 76% of control . In a few days following the implementation
of a strong restriction of access time to drink (½h/24h),
rabbits have a very strong reduction in the consumption of solid
food and water (figure 29). The reduction in the time devoted
to the consumption of solid food is reduced in the same proportions,
suggesting a constant rate of ingestion. Rabbits gradually adapt
and stabilize their feed consumption in just over 8 days depending
on the age of the animals and of the actual time of access to
water.
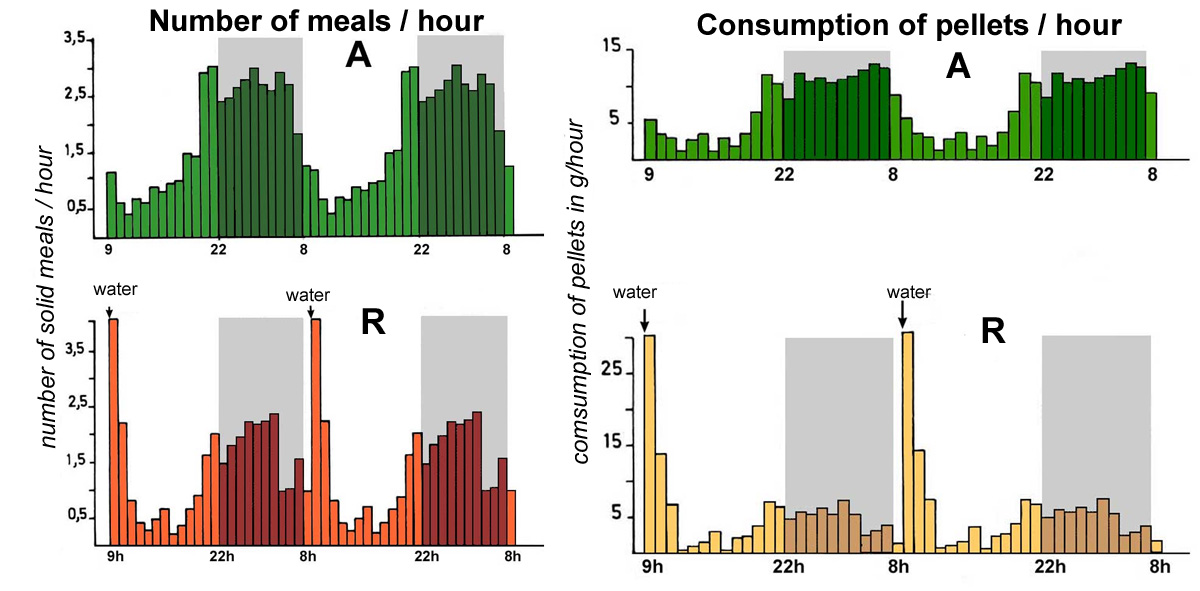
After some
times of adapatation (one or two weeks) rabbits authorized to
drink water only for a limited time per 24-hour cycle, increase
the frequency of their solid meals immediately after the distribution
of water, (4 solid meals per hour compared to 1 only for the
control rabbits). The amount of pelleted feed consumed per hour
follows the same profile with a peak immediately after the water
distribution (figure 30). After this initial peak of consumption,
rabbits then have an hourly feed consumption profile quite similar
to that of rabbits with permanently available water but with
lower quantities
|
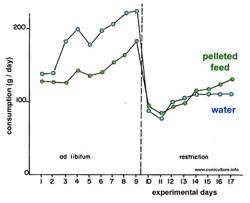 Figure
29: Daily water and feed consumption before and after a
restrition of access to drinking water of ½h per 24h.
Figure
29: Daily water and feed consumption before and after a
restrition of access to drinking water of ½h per 24h.
(Source: Lebas and
Delaveau, 1975). |
|
|
|
Figure
30 : Evolution over 48 hours of the number of solid meals
per hour and of the quantity of pelleted feed consumed each
hour in rabbits reveiving water ad libitum (A in
green) or receiving water during 10 min /day at 9h00 ( B in
yellow-beige). The experimental building
was lit from 8h00 to 22h00. Source:
Prud'hon et al. (1975).
|
|
| |
|
|
|
Under practical
conditions, access to water limited to a period of 1h30 to 4h
leads to a more marked reduction in water ingestion than the
induced reduction in feed consumption, in particular for the
shorter periods (Figures 31 and 32). This difference in behaviour
results in a reduction in the water / food ratio from 1.74 in
rabbits fed ad libitum to 1.54 in those receiving water
only for 1 hour 30 minutes or 2 hours per 24 hours.
It should
be noted that in case of water restriction, the water / food
ratio is always reduced due to a reduction in water intake more
marked than that of solid feed. On the other hand, a reduction
in the quantity of feed daily allocated with the same magnitude,
or even more important, the ingestion of water is on the contrary
greatly increased and exceeds that of control animals fed ad
libitum. As a result, the water / feed ratio is greatly
increased (table 13).
|
|
Intake
|
Ad
libitum
(control = 100)
|
|
Feed
65%
|
|
Pellets
|
136
g/d = 100
|
78
|
65
|
|
Water
|
228
g/d = 100
|
56
|
136
|
|
water/feed
|
1.7
|
1.2
|
3.5
|
Table 13
: Effects
of limiting the daily duration of access to drinking water or
of a reduction in the quantity of pelleted feed distributed every
day, on the relative consumption of water and feed.
Source: Boisot et al., (2005). Averages per 24h during
the 3 weeks following weaning at 31 days.
|
|
|
|
|
Figure
31 : Water consumption in function of access to water
|
Figure
32 : Feed consumption in function of access to water
|
|
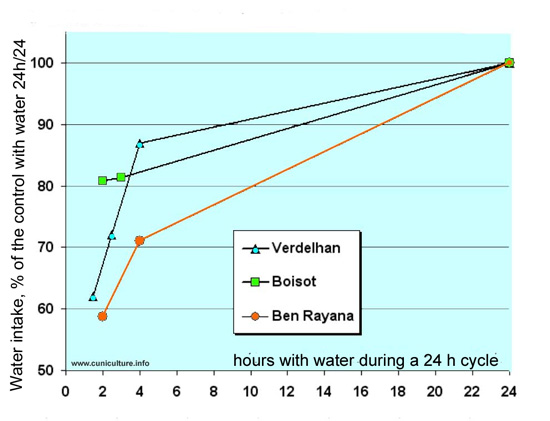
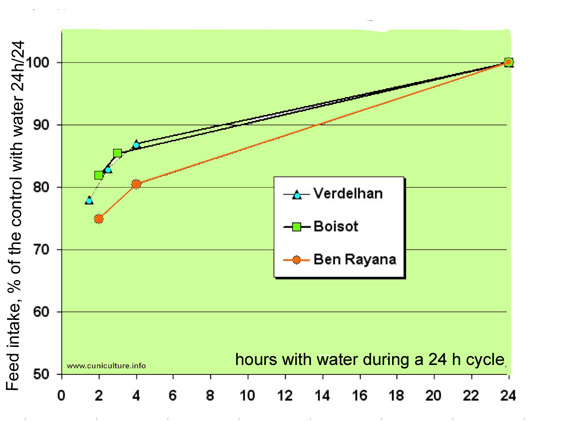
Relative
consumption of rabbits with a access to water only 1.5 to 4
hours per 24h or continuously, but always fed ad libitum
Data of 3 authors,
Verdelhan et al. (2004), Boisot et al. (2004) and Ben Rayana
et al. (2008). The results are expressed as a percentage of
the control groups with a continuous access to water and to
pelleted feed.
|
|
| |
|
|
Feed
restriction and importance of the digestive tract |
| |
|
|
As
a general rule, any rationing less than 80-85% also tends to reduce the
growth rate of the animals. On the other hand, the digestive tract is
significantly less affected by feed restriction than the body as a whole,
as shown by the data in Table 14. Moreover, if a food restriction systematically
promotes an increase in digestive content, the distribution of the latter
between the different segments depends largely on the restriction mode
selected. |
| |
|
|
| |
Restriction
Mode
|
| |
Ad
Libitum |
Quantitative
restriction (1) |
| Distibution
periodicity |
24h
/ 24
(control)
|
5d
/ 7
(= 71% of the time) |
Daily distribution
of 71% of control
|
Distribution
of 71%
twice /week
|
|
Observed
consumption
(% of the control group)
|
100% |
78.9% |
70.4% |
71.4% |
|
Slaughter weight (g)
|
2016 |
1918 |
1901 |
1875 |
|
Average
Daily Gain (g / d)
|
38.1
a |
21.3
c |
25.7
b |
16.6
d |
|
Empty
digestive tract (g)
|
149
a |
142
b |
142
b |
141
b |
|
Digestve
tract content (g)
|
220
a |
273
b |
329
c |
316
c |
|
Digestive content found in the different segments expressed
as % of the total digestive content
|
|
-
Stomach
|
37.3
a |
33.8
c |
40.9
b |
39.1
b |
|
-
Small intestine
|
11.8
a |
11.5
a |
15.0
b |
12.2
a |
|
-
Caecum
|
40.4
a |
45.2
b |
35.8
c |
40.8
a |
|
-
Colon
|
10.6
a |
9.8
a |
8.4
b |
7.9
b |
(1) Restriction
calculated according to a previous experiment with the same pelleted
diet
Table 14 :
Incidence of various modes of feed restriction on digestive development
in rabbits slaughtered on average at 67 days.
Source: Lebas
and Laplace (1982). [Commercial complete pelleted feed containing 16.5%
proteins and 14.0% crude fiber].
|
| |
|
|
|
Due to the
greater relative development of the digestive mass and especially
of its content, the different modes of restriction significantly
affect the slaughter yield, but the effect is directly related
to the speed of growth as shown by the results of the figure
33.
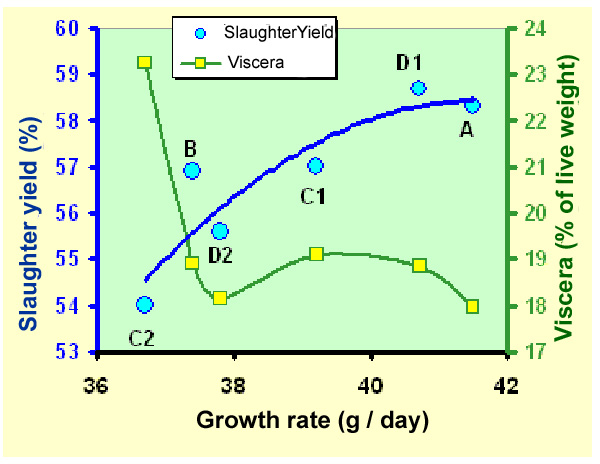
Figure
33. Slaughter yield (blue dots) and proportion
of full viscera (green-yellow squares) in young rabbits
slaughtered at 70 days, in function of the growth rate observed
during the fattening period, using different types of feed restriction.
Source: Jerôme
et al. (1998) . All the rabbits were fed again ad libitum one
week before slaughter.
A=
control group fed ad libitum continuously
B=
80 % quantitative restriction, with daily distribution
C1=
access to the feeder limited to 16 h / 24h., at night
C2=
access to the feeder limited to 8 h / 24 h., during the lit
period
D1=
access to the feeder prohibited 2 times 8 hours each week, otherwise
fed ad libitum
D2=
access to the feeder prohibited for 24 h., once a week, otherwise
fed ad libitum
|
|
|
| |
|
|
From
a practical standpoint, it should be remembered that any restriction,
reducing the growth rate, will also reduce the slaughter yield .
|
| |
Feed
preferences in rabbits |
| |
|
|
Feeding
behavior of wild non-captive rabbits ("grazing" rabbits) |
| |
|
|
|
| |
|
|
|
| |
|
|
|
The wild rabbit
can feed on a very wide range of plants. However, it clearly prefers
graminaceous (Fescue, Brachypodium, or Digitaria for example)
and consumes few dicotyledons if other plants are available. Among
the dicotyledons it prefers certain leguminosae (Fabaceae, Mimosaceae,
Papillonaceae, ...) and asteracea (formerly Compositae). But it
should be noted that if rabbit have the choice, the consumption
of carrots (Daucus carotta) is very low. Contrary to popular
belief, carrots (roots or leaves) are not among the plants sought
after by rabbits.
The proportion
of dicotyledons and even mosses can increase during some seasons
when plant availability is low. In winter and early spring, grazing
by rabbits of young cereals grown can completely compromise a
harvest, especially in the 30 to 100 m area from the burrow. When
rabbits can choose between winter cereals grown with or without
mineral fertilization (phosphorus and / or nitrogen) they clearly
prefer cereals without artificial fertilization.
In a situation
of choice, the rabbit can be very selective in its feeding behavior.
Like many herbivores, it will prefer to ingest the leaves rather
than the stem of a plant, and generally young plants or "green
and tender" parts rather than dry ones. For example, he will
choose a part of the plant with a high nitrogen concentration.
Likewise, in a trial conducted in Ireland, rabbits grazed more
intensively a particular variety of spring barley compared to
4 other winter barley varieties. This is possibly related to the
composition of the plant. However, the differences in the sugar
content of the 5 varieties did not fully explain this varietal
choice.
At the end
of winter, the rabbit has a strong appetite for the buds and young
stems of some woody plants. The browsing of very young trees or
their shoots can thus completely compromise the regeneration of
a forest, or more specifically the regeneration of certain shrubs
such as juniper or the Scotch broom (Cytisus scoparius)
as is observed in France in Sologne. In winter the rabbit likes
to eat the bark of some cultivated trees (not just the young stems),
especially apple trees and also peach or cherry trees. The bark
of apricot, pear and plum trees is generally less attacked.
|
|
|
In the forest,
rabbits clearly prefer deciduous trees, but can also attack the
bark of conifers (mainly firs and some types of pines). When the
trees are very young, rabbits prefer to eat the apical or lateral
shoots of fir trees, rather than those of oaks.
The basic
reasons for the rabbit's feed choices remain unclear, even if
they are constant. It can only be said that this behavior is under
hypothalamic regulation, since lesions of the hypothalamus clearly
modify the rabbit's feed choices.
Many experiments
have been undertaken particularly in Australia and New Zealand
to study the behavior of wild rabbits, with a view to develop
baits (the end goal being the extermination of imported wild rabbits
in the 19th century). It was observed a lot of variation depending
on the type of bait, but also depending on the season. For example,
pellets made from milling by-products (middlings + bran) are well
consumed throughout the year. In contrast, the acceptability of
carrots or oats changes seasonally. The addition of salt (1% or
5% NaCl) or alfalfa meal (15%) in the bait pellets made with wheat
by-products significantly reduces their consumption.
|
 Rabbit damages
on a young pine tree
Rabbit damages
on a young pine tree |
|
| |
|
|
Feeding
behavior of domestic rabbits in a free choice situation (rabbit in
cage with a choice for their feed) |
| |
|
|
Some
studies have shown that the rabbit can recognize basic flavors, such as
salty, sweet, bitter, sour. He shows a preference for sweet flavors, and
chooses for example a feed containing additional sugar or molasses rather
than a food of the same composition containing no additional sweeter. |
| |
|
|
In
captivity, the adult rabbit can sometimes express a "delicate"
feeding behavior, with a momentary refusal of ingestion after a change
of feed, or a systematic refusal of certain feeds. It is then observed
a scratching behavior of the content of the feeder, whether the feed is
in pelleted form or in flour. |
| |
|
|
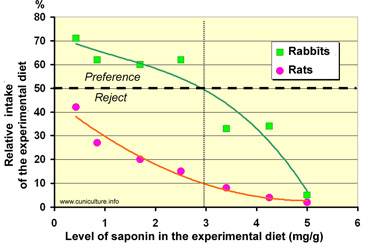 Figure 34
: Test of free choice consumption of rabbits or rats, between
a feed without saponin and a feed containing various levels of
saponin (source of bitterness) provided by alfalfa.
Figure 34
: Test of free choice consumption of rabbits or rats, between
a feed without saponin and a feed containing various levels of
saponin (source of bitterness) provided by alfalfa.
Source: Cheeke (1974).
|
If a rabbit
can choose between a feed with or without an appetite stimulator,
it usually chooses the feed with the "apetizer". But,
if each of these 2 feeds is offered alone, there is no difference
in ingestion, or in growth. The same phenomenon has also been
shown with the addition of a repellent in the feed such as formalin.
On the other
hand, the rabbit seems to appreciate a certain degree of bitterness
in its diet. When rabbits are presented with feeds containing
dehydrated alfalfa with variable levels of saponin, and therefore
variable bitterness, their choice is fixed on those feeds which
have a relatively high degree of bitterness (up to 3 mg/ g of
saponin in the diet - figure 34). All these feeds are neglected
for example by rats or pigs. On the other hand, if only feeds
with a variable rate of saponin (1.8 to 6.4 mg / g of feed) are
proposed, feed intake and growth are independent of the saponin
concentration.
When the diet
contains a toxin, such as aflatoxin, the rabbit totally refuses
to consume it, or he ingests it in very small quantities. When
an energetic feed (low in fiber) is distributed with a free choice
with a fibrous feed, the rabbit generally prefers the first. It
is probably the consequence of a specific search for energy sources
(rare in nature), which is the regulatory system dominating the
ingestion of the rabbit. On the other hand, for the rabbit, this
can lead to an increase in the frequency of digestive disorders
(lack of fibre and consequently slow digestive transit) and therefore
to enhanced health risk, particularly in growing young.
|
When a rabbit is offered
different "normal" types of feeds, its choice for one or the
other is often unpredictable. When offered ad libitum dehydrated
alfalfa pellets and dry maize grains, the proportion is 65% alfalfa and
35% maize. In the case of alfalfa and oats the proportion is 60/40. However,
if the grains of maize are relatively humid (over 14-15 %, which may cause
storage problems), the proportion of grain increases to 45-50 %. |
| |
|
|
Feeding
rabbits on roughages + a complementary concentrated feed may cause problems
if the roughage does not taste good. When rabbits are free to choose between
concentrated energy feed and fibre (straw, for example), they are incapable
of balancing the ingestion of both feeds and growth decreases. If producers
find themself in this situation, they must limit the amount of concentrated
feed, or the feed that the rabbits prefer. This is what happens in the
case of certain green roughages of little nutritional value. As observed
by Gidenne (1985), the situation is quite different if the rabbit is offered
two concentrated energy feeds, such as complete pelleted feed and green
bananas. The growth rate of the rabbits with free choice is similar to
that of the control (only complete pellets), and the ingestion of digestible
energy is identical to that observed with only pelleted feed. In any case
between weaning (5 weeks) and the end of thel (12 weeks), the proportion
of bananas consumed dropped from 40% down to 28% of the daily intake of
dry matter. |
| |
|
|
Finally, it
should be noted that growing rabbits, which receive a pelleted feed
deficient in sulfur amino acids or in lysine, and which simultaneously
have a free choice between pure water and for drink a solution of
the deficient amino acid, the amino acid solution is clearly preferred
to pure water (table 15).
They thus succeed
in having a growth similar to that of the control rabbits receiving
a balanced diet. It should also be noted that in this test the drinking
trough receiving the pure water solution and that receiving the
amino acid solution were alternated each day. While some rabbits
did indeed change the "preferred" drinker depending on
its content, other rabbits did not change their "favorite"
drinker but drank little or a lot, depending on the nature of its
content, which corresponds to two types of behavioral adaptation.
|
|
Amino-acid
in water
|
Préference
for water with the amino acid
|
|
Lysine
1.6 /Liter
|
56%
|
|
|
77%
|
|
Methionine
3g/L
|
60%
|
Table 15
: Preference rate of growing rabbits fed a feed deficient in
an amino acid and optionally as drink pure water and a solution
of the deficient amino acid in the feed.
Source:
Lebas and Greppi (1980).
|
|
| |
|
|
|