| |
| Entéropathie
épizootique du Lapin (EEL) |
|
Compte tenu
de l'importance du sujet et de la préoccupation des éleveurs,
au moins en Europe, on peut s'étonner du petit nombre de
communications faites sur l'EEL (3 + la présentation de synthèse).
Il est vrai que lors de la table ronde sur les entéropathies
du lapin, animée par Badiola (Espagne), une grande partie
de la discussion a été dévolue à cette
pathologie.
Dans son rapport de synthèse D. LICOIS [12] a fait le point
sur les dernières avancées obtenues depuis le congrès
de Valence (2000). L'obtention d'un inoculum de référence
(TEC), a permis de caractériser précisément
l'EEL (cinétique de la maladie, signes cliniques, lésions
macroscopiques…) et de confirmer l'absence de spécificité,
voire l'absence tout court, des lésions histologiques, qui
s'avèrent donc inutilisables en terme de diagnostic. Avec
l'appui de différents partenaires, la caractérisation
de l'inoculum au niveau parasitaire, bactériologique et virologique
a permis d'écarter un certain nombre d'hypothèses
étiologiques.
|
| |
En ce qui concerne
l'étiologie, bien que des rotavirus aient été
identifiées, ils ne sont probablement pas les agents primaires
responsables de la maladie. L'autre germe retrouvé est Clostridium
pefringens. Une coopération avec une équipe belge
(D. Marlier et H. Vindevogel) ciblée sur le rôle de
cette bactérie et de ses toxines alpha et béta2, a
été engagée mais aucun résultat univoque
n'est venu étayer cette hypothèse pour le moment.
De même, une firme privée (Proteus) a développé
un programme d'identification de l'agent pathogène de l'EEL,
par différentes approches de biologie moléculaire.
Des pistes sur l'intervention possible de clostridiales d'une part
ou d'une famille de virus d'autre part, ont été énoncées
mais sans résultats tangibles ultérieurs. Néanmoins
lors de la table ronde, Badiola a mentionné que sur 4 souches
de Clostridium pefringens isolées de lapins atteints
d'EEL, l'une d'entre elles avait reproduit des symptômes d'EEL
et entraîné jusqu'à 50% de mortalité.
Une autre bactérie dont le nom n'a pas été
cité, pourrait également intervenir dans l'étiologie
de l'EEL. Il serait particulièrement intéressant de
voir comment se comportent ces germes dans nos conditions expérimentales
(lapins EOPS - Exempts d'Organisme Pathogènes
Spécifiés) et de vérifier si ces bactéries
sont présentes dans notre inoculum ou dans ceux obtenus par
d'autres équipes.
|

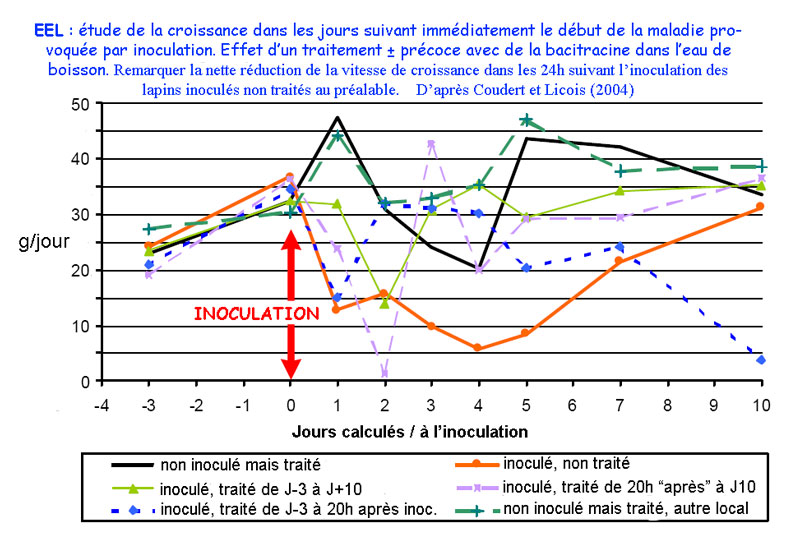
Evolution du gain de poids de lapins après
inoculation de l'EEL chez des lapins traités ou non avec
de la Bacitracine

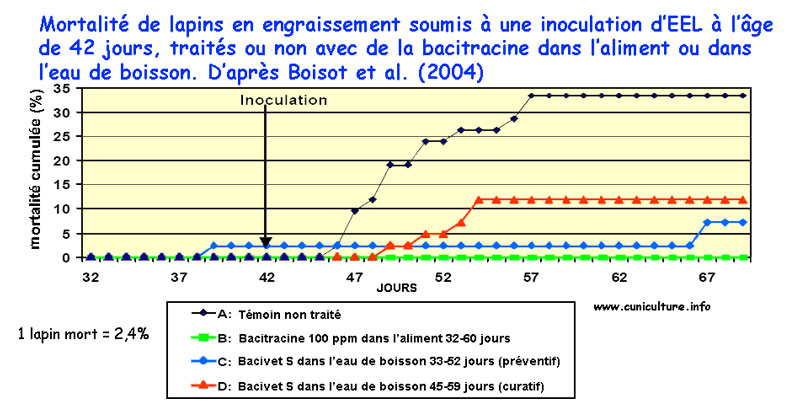
Mortalité
cumulée chez des lapins en engraissement soumis à
une EEL provoquée, traités ou non avec de la Bacitracine
mise dans l'eau ou l'aliment.
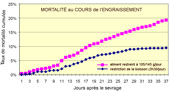
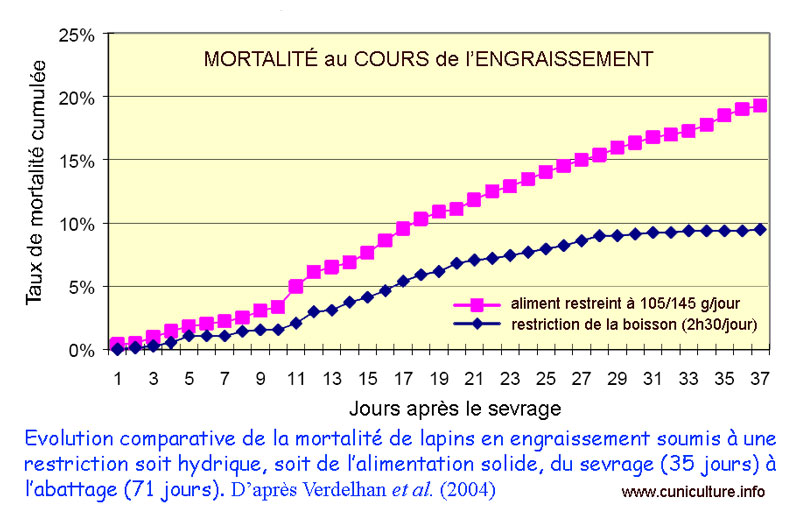
Mortalité
cumulée de lapins rationnés via l'aliment (violet)
ou via l'eau de boisson (bleu)
|
Des indications
concernant aussi l'étiologie ont été présentées
par COUDERT et LICOIS [10] qui ont analysé
les évènements précoces survenant lors de reproductions
expérimentales de l'EEL. Une chute significative mais transitoire
du gain de poids est observée moins de 20h après l'inoculation
des animaux, y compris chez ceux traités au Bacivet S et
qui n'ont pas développé d'EEL. Ces auteurs suspectent
l'intervention possible d'une toxine présente dans l'inoculum
pour expliquer ce phénomène.
L'action de la bacitracine soluble dans l'eau de boisson (Bacivet
S®) a été comparée à celle préalablement
utilisée en supplémentation dans l'aliment (Albac®),
chez des lapins conventionnels inoculés avec l'inoculum TEC3
[6]. Le Bacivet S (à 0.675 g/l) donne d'aussi bons résultats
que l'Albac à 100 ppm, si l'utilisation est faite en préventif
: réduction significative de la morbidité et de la
mortalité comparativement aux animaux inoculés non
traités. Par contre, en curatif le traitement au Bacivet
S, bien que réduisant aussi morbidité et mortalité,
est moins efficace.
Soulignons un autre travail qui n'a pas été réalisé
dans le cadre strict de l'EEL mais dont l'application parait tout
aussi utile pour cette pathologie. Il s'agit de la restriction hydrique [16]. Il avait déjà été montré
les effets bénéfiques d'une restriction alimentaire
(entre 60 et 80%) de l'ad libitum sur la réduction
de la mortalité et de la morbidité, en situation d'EEL
ou non. Ici, l'étude a été réalisée
dans un élevage qu'on peut supposer comme ayant des problèmes
digestifs. La méthode proposée a consisté à
laisser l'accès à l'eau de boisson pendant un courte
période de la journée (ici 2h30) ce qui a induit indirectement
une restriction alimentaire de l'ordre de 83% de l'ad libitum.
Comparativement à une restriction alimentaire vraie, la restriction
hydrique a réduit la mortalité (9.3% vs 19.3%).
Dommage qu'il n'y ait pas eu de lot témoin qui aurait pu
préciser le taux de mortalité en l'absence de restriction
hydrique ou alimentaire, ni d'analyse de la morbidité (GMQ
par exemple).
|
| |
Enfin dans la
session génétique, DE ROCHAMBEAU et al. [14] ont rapporté des résultats concernant un programme
sur la résistance génétique aux entéropathies
du lapin, incluant l'EEL. Dans cette communication a été
analysée la variabilité génétique de
lapereaux, issus de 48 mâles, soumis à trois modèles
de pathologie digestive expérimentale (coccidiose, EEL et
pathologie non spécifique provoquée par une réduction
du taux de fibres alimentaires). Plusieurs indices ont permis de
caractériser la réponse individuelle des animaux.
Pour chaque modèle un effet significatif des mâles
sur certains indices a été démontré.
Ces résultats montrent l'existence d'une variabilité
génétique pour la résistance aux 3 modèles
étudiés. Signalons que des travaux sont poursuivis
actuellement plus spécifiquement sur l'EEL.
|
| Escherichia
coli et colibacilloses |
Les colibacilloses
demeurent avec l'EEL l'une des pathologies dominantes actuellement
en élevage. Essentiellement dus aux Escherichia coli
appartenant à la catégorie des EPEC (enteropathogenic
E. coli), certains pathovars sont responsables de pertes
importantes après sevrage mais parfois aussi avant sevrage.
Plusieurs auteurs ont rappelé les avancées obtenues
ces dernières années sur la connaissance des mécanismes
de pathogénicité et l'identification des facteurs
de virulence et l'utilisation qui pouvait en être faite en
termes de diagnostic, d'épidémiologie ou de prophylaxie
vaccinale.
En plus du rapport
introductif de LICOIS [12], 11 communications sur les 44 de la session
pathologie font référence aux E. coli intestinaux
(soit 27%), c'est dire l'importance de cette bactérie, au
moins en termes d'objet de recherche ou d'étude. C'est en
tout cas nettement plus que les quelques communications produites
dans ce domaine lors des 2 précédents congrès
(Valence : 3, Toulouse : 2).
|
| |
Les
communications ont été regroupées en 3 catégories
- celles qui s'intéressent au diagnostic
et à l'épidémiologie
- celles qui traitent de vaccination à
partir de souches mutées
- et les diverses : enquêtes terrain,
abattoir, traitements…
|

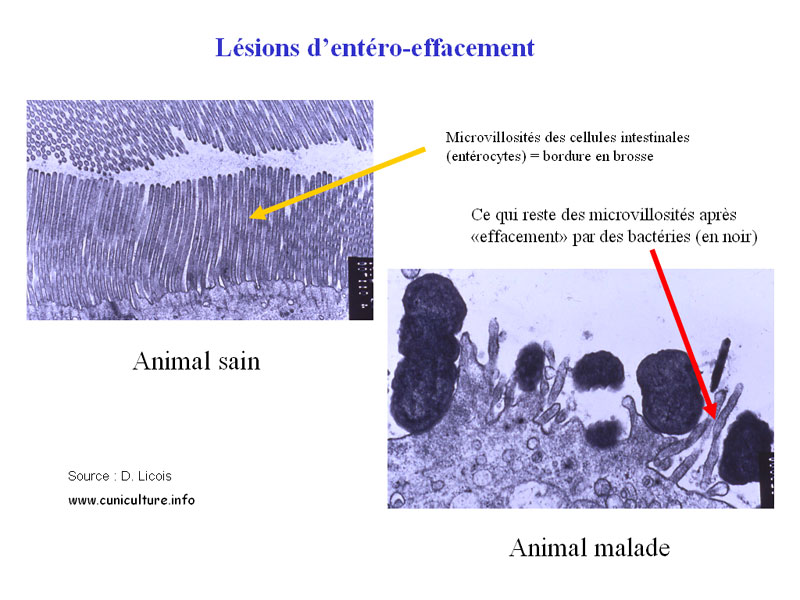
Illustration histologique d'une lésion d'effacement
|
Rappel sur
la pathogénicité, les facteurs de virulence et les
mécanismes moléculaires mis en jeu [12]
Les EPEC ainsi
que les EHEC (enterohemorrhagic E. coli) induisent des lésions
spécifiques d'attachement-effacement à la surface
des cellules épithéliales intestinales de l'iléon
du cæcum et du côlon qui sont les segments intestinaux
concernés par les EPEC chez le lapin. Ces lésions
sont caractérisées par un attachement intime des bactéries
au niveau de la bordure en brosse des entérocytes et par
la formation de structures appelées "piedestals"
dans lesquels sont enchâssées les bactéries.
Les connaissances acquises depuis le début des années
1990 sur les facteurs de virulence des EPEC résultent des
travaux réalisés in vivo et in vitro
(cultures cellulaires) à partir de souches entéropathogènes
du lapin (Milon, Peeters, Blanco…) ou d'une souche EPEC humaine
de référence.
De manière synthétique un modèle en trois étapes
a été proposé.
La première étape correspond à une adhésion
modérée médiée par des protéines
fimbriales, facteurs d'attachement codés par des plasmides
et appelées adhésines. Elles permettent aux bactéries
de coloniser le tractus intestinal et de s'opposer aux mécanismes
de résistance non spécifiques comme le péristaltisme.
Chez le lapin, deux adhésines ont été décrites
: la première AF/R1 chez une souche O-15 et la seconde AF/R2
chez les souches EPEC O-103.
Le deuxième et le troisième stades conduisent
à un contact étroit entre la bactérie et la
cellule cible, l'enterocyte. Ils impliquent plusieurs protéines
codées par des gènes chromosomiques responsables des
lésions d'attachement effacement, localisés dans un
îlot de pathogénicité le Locus d'Entéro
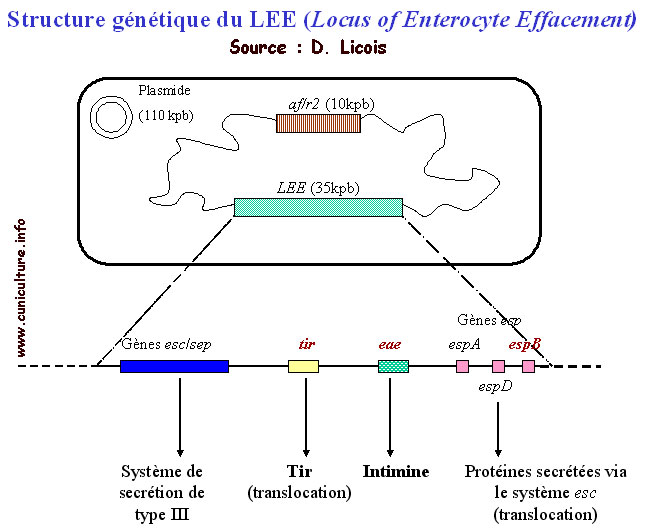
Effacement (LEE). Le LEE code pour différentes protéines
ayant une grande diversité de fonctions. La plupart d'entre
elles sont secrétées via un système de sécrétion
de type III (TTSS). Dans la région centrale du LEE, le gène
eae
code pour une protéine de membrane externe, l'intimine qui
se fixe sur son récepteur Tir
(translocated intimin receptor) et établit ainsi un contact
étroit entre la bactérie et l'entérocyte. Tir
est secrété par le TTSS et transloqué (injecté
dans la cellule hôte) à travers la membrane cellulaire
par l'intermédiaire d'une "seringue moléculaire"
qui comprend plusieurs effecteurs, EspA, EspB and EspD, (E. coli
secreted proteins). Toutes ces protéines sont nécessaires
pour activer un signal qui conduira aux lésions d'attachement-effacement
et aux perturbations physiopathologiques associées à
la diarrhée.
|
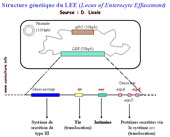
Structure génétique du LEE (Locus d'Entéro Effacement)
|
Constructions
de souches vaccinales et essais de protection
BOULLIER et al. [7] ont obtenu à partir d'une souche
sauvage d'E. coli O103 Rh- virulente appelée E22, une
souche dont 2 gènes du LEE ont été rendus non
fonctionnels : les gènes EspB et Tir. Ces auteurs
ont ensuite vérifié que cette souche mutée (E22 D Tir / EspB) n'était pas pathogène pour le
lapin (pas d'induction de diarrhée ni de lésions histologiques)
mais conservait une capacité à se multiplier chez l'animal
(ce dernier point est important pour que le contact bactérie-hôte
puisse entraîner une réponse immunitaire et une protection
efficace). Il a également été démontré
que les animaux vaccinés avec la souche vaccinale étaient
protégés contre une inoculation d'épreuve avec
la souche virulente E22, aussi bien 8 jours que 28 jours après
vaccination. De plus la souche mutée réduit l'excrétion
de la souche sauvage et donc le risque de dissémination de
cette souche virulente entre animaux. Enfin des anticorps dirigés
contre le LPS O-103, l'intimine et l'adhésine AF/R2 ont été
détectés dès le 7e jour suivant la vaccination. La
conclusion de ce travail est que la souche mutée E22 D Tir/EspB est un bon candidat vaccin contre la colibacillose O-103. La question
qui reste posée est de savoir s'il existe une protection croisée
vis-à-vis d'autres pathovars EPEC du lapin (O-128, O-15, O-132,
O-109…) et si oui à quel degré.
|
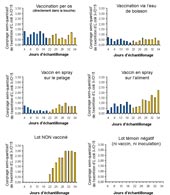
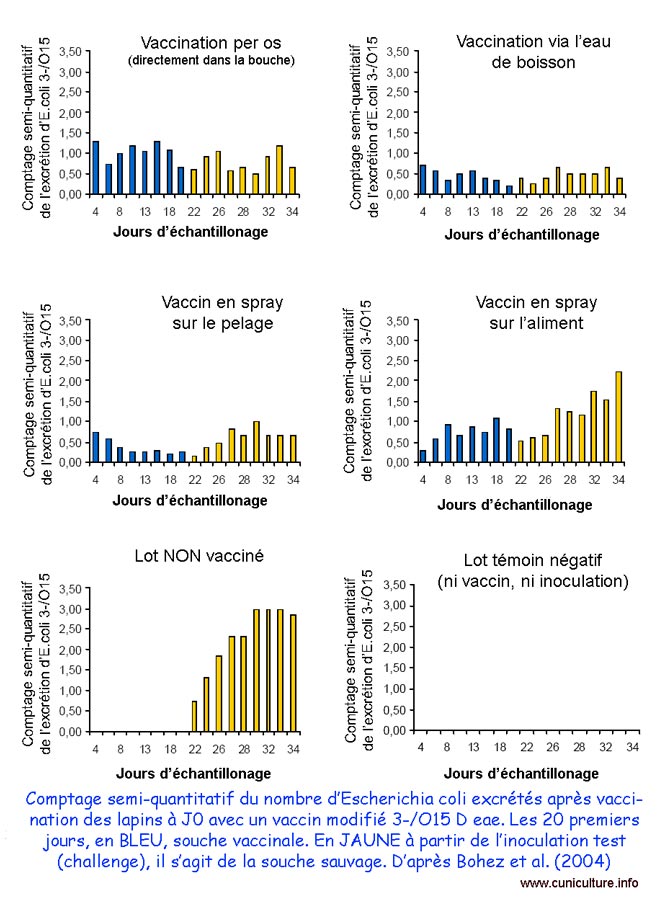
Comptage des
E. coli avant et après vaccination avec une souche
modifiée
(O15 D eae)
administrées par différentes voies |
C'est ce qu'ont essayé d'évaluer Bohez et al. [3, 4, 5] qui ont travaillé sur deux autres souches entéropathogènes du lapin : une souche d'E. coli O-15 délétée dans le gène eae et une souche O-132 délétée dans le gène tir. Rappelons que les souches O-15 dominent en Belgique, alors qu'en France ou en Italie les souches O-103 sont les plus fréquentes.
La souche mutée O-15 D eae a été utilisée en condition de terrain dans un élevage où sévissaient des problèmes de colibacillose associés à l'entéropathie épizootique [3] . En administrant la souche vaccin à des lapins de certaines cages, les auteurs ont d'abord cherché à savoir si cette souche était capable de diffuser aux animaux non vaccinés des cages voisines. Le résultat a été négatif. La conclusion est qu'il est nécessaire de vacciner l'ensemble des animaux. Le deuxième objectif était de vérifier l'efficacité de la vaccination. Aucune différence n'a été montrée entre le lot vacciné et le lot non vacciné en ce qui concerne les paramètres zootechniques et cliniques enregistrés (mortalité moyenne 20%, morbidité moyenne 26%). Dans les deux lots ont été retrouvés les sérogroupes O-109 (EPEC rencontrés chez les lapereaux avant sevrage) et O-132. Ceci rejoint les résultats obtenus en 2003 par cette équipe, avec la même souche O-15 D eae, mais en condition expérimentale cette fois. Elle avait démontré que cette souche protégeait bien vis-à-vis de souches sauvages homologues O-15 alors qu'elle confirme ici que la protection contre des souches hétérologues O-132 ou O-109 n'est pas totalement satisfaisante.
Dans une autre présentation, intéressante sur le plan pratique, ces mêmes auteurs [5] ont testé différentes modalités d'administration de la souche atténuée O-15 D eae (per os, via l'eau de boisson, par pulvérisation sur aliment ou directement sur le poil des animaux). Ils ont ensuite vérifié expérimentalement son efficacité par une inoculation d'épreuve avec une souche sauvage O-15. Dans tous les cas de figures la souche vaccin s'implante bien et diminue l'excrétion de la souche sauvage après challenge. Par contre la virulence modérée de la souche sauvage utilisée dans cet essai ne permet pas d'apprécier l'efficacité réelle de la vaccination sur l'expression de la pathologie.
|
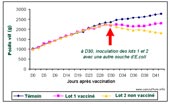
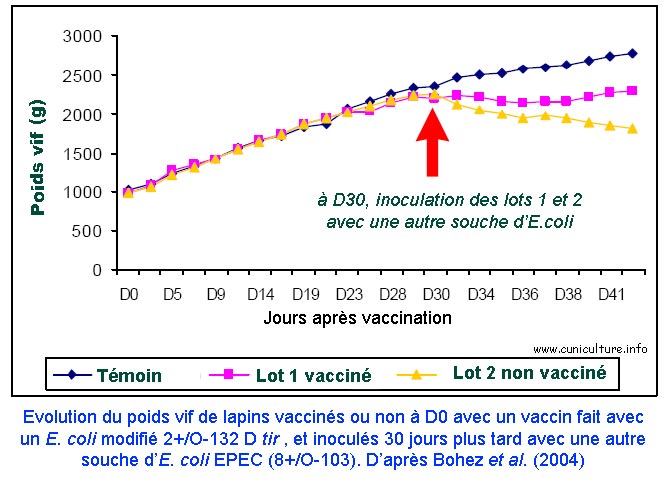
Evolution du
poids vif de lapins vaccinés avec une souche d'E.
coli atténuée (O132 D tir) et inoculés
30 jours après avec une souche sauvage de serotype O103. |
La deuxième souche atténuée O-132 D tir, étudiée par Bohez et al. [4] , dérive d'une souche sauvage O-132, considérée comme moins pathogène que les E. coli O-103 ou O-15. L'objectif était précisément ici de tester expérimentalement la protection croisée de cette souche vaccinale vis-à-vis de souches virulentes hétérologues O-15 et O-103. Les résultats obtenus montrent que la protection a été bonne contre la souche O-15 mais insuffisante vis-à-vis de la souche O-103.
La conclusion générale sur ces souches mutées est qu'expérimentalement on a de bons résultats de protection face aux pathotypes dont sont issues directement les souches vaccinales, mais il reste à trouver un vaccin polyvalent qu'il faudra aussi valider sur le terrain (pourquoi pas une association de différentes constructions comme le propose Bohez et al. [4] ).
|

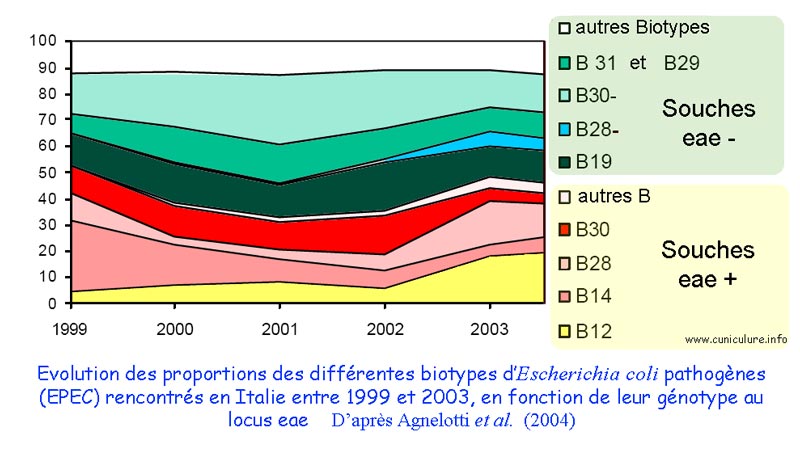
Evolution des
biotypes d'E.coli dans les élevages cunicoles italiens
entre 1999 et 2003, avec identification du gène eae. |
Diagnostic et épidémiologie
Ce domaine a été abordé dans quatre communications émanant essentiellement d'auteurs italiens (Agnoletti et al. [1] , Camarda et al. [8] , D'Incau et al . [11] , Pisoni et al. [13] ) . Tous se sont intéressés à caractériser des souches d' E. coli isolées de lapins atteints de troubles digestifs. Les critères utilisés sont le sérogroupage (identification de l'antigène somatique O), biotypage (aptitude à fermenter certains sucres), profil d'antibiorésistance et certains facteurs de virulence : recherche du gène eae ou de gènes codant pour des toxines. Là aussi les avancées de la biologie moléculaire ont été utilisées. La plupart des auteurs ont inclus le gène eae comme outil de diagnostic pour identifier les souches virulentes afin d'évaluer la prévalence des colibacilloses sur le terrain, dans différentes régions de l'Italie. Je citerai un seul exemple, représentatif a mon sens, puisqu'il concerne plus de 2000 souches d' E. coli isolées d'élevages intensifs de l'ensemble de l'Italie et analysées pour le gène eae [1] : selon ces auteurs, la prévalence des EPEC avait régressé en Italie entre 1999 et 2000, pour s'accroître depuis, avec 45% des souches possédant le gène eae. Cependant il faut rester prudent car des travaux français réalisés il y a une dizaine d'années avaient montré que le gène eae pouvait être identifié chez des souches O-103 non pathogènes. Les sérogroupes les plus fréquemment rencontrés sont O-103 et O-2 [11] . Enfin, alors que les souches isolées possèdent de nombreux profils d'antibiorésistance, la sensibilité aux quinolones (notamment la marbofloxacine) reste importante [8, 13] .
|
|
|
Autres travaux
Badiola et al. [2] se sont appuyés sur des techniques utilisées en biologie moléculaire notamment la RFLP (restriction fragment length polymorphysm) pour différencier des profils de flores microbiennes chez des lapins malades comparativement à des lapins sains. En utilisant 5 enzymes de restriction appliquées à un fragment du gène ribosomal bactérien 16S, amplifié par PCR à partir de contenu intestinal, ces auteurs ont observé des profils RFLP caractéristiques de disfonctionnements intestinaux associés à l'entéropathie épizootique, aux colibacilloses, aux clostridioses ou même à des modifications liées à l'antibiothérapie. Ces profils sont ensuite introduits dans une base de données conçue et gérée par les auteurs. L'intérêt de ce type d'approche est multiple : il permet de s'intéresser à des bactéries non cultivables, de mieux appréhender des écosystèmes complexes comme celui de la flore microbienne intestinale, et enfin d'utiliser la base de donnée comme outils d'aide au diagnostic.
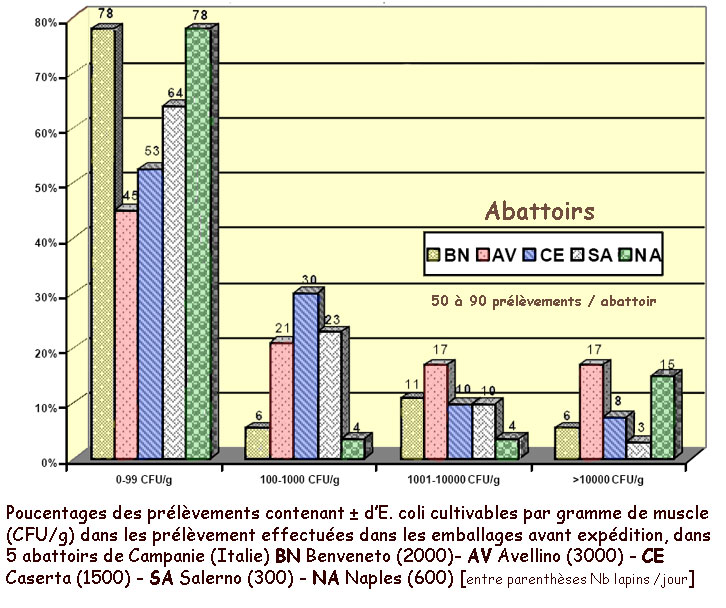
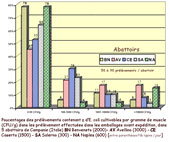
Le papier de
Cerone et al. [9] avait pour objet la
recherche d'agents principalement bactériens responsables
potentiels de zoonoses (Salmonelles, Lysteria, Campylobacter,
.),
au niveau des 5 abattoirs de la région de la Campanie en
Italie. E. coli, entre autres, a été inclus
comme marqueur des qualités hygiéniques. En effet
cette bactérie, essentiellement présente dans l'intestin,
peut se retrouver sur les carcasses, voire dans l'environnement
après éviscération des animaux. Elle peut donc
constituer un bon marqueur des pratiques utilisées, et donc
de l'hygiène, tout au long de la chaîne d'abattage.
Les auteurs ont effectivement trouvé des abattoirs où
une proportion élevée de prélèvements
contenait beaucoup [trop] d'E. coli : plus de 10 000 /g (voir
figure ci-contre)
Un dernier travail présenté par Tsalie et al. [15] a consisté à évaluer l'incidence de la vitamine E sur les caractères morphométriques des villosités intestinales de l'iléon ainsi que sur les leucocytes intraépithéliaux, lors d'une colibacillose expérimentale (souche O-103 Rh- E22). La vitamine E est connue pour stimuler la réponse immunitaire dans un certain nombre d'affections. L'inoculation des colibacilles seule entraîne une réduction de l'épaisseur de la muqueuse et de la hauteur des villosités A l'inverse, une amélioration significative de ces paramètres est observée lorsque l'inoculation est associée à l'administration de vitamine E. De même les mononucléaires et les polynucléaires de la lamina propria sont plus élevés chez les lapins inoculés recevant la vitamine E par rapport aux animaux inoculés non traités, ce qui rend compte d'une réaction inflammatoire plus intense. Par contre, il n'a pas pu être mis en évidence de différence significative entre les 2 groupes d'animaux, concernant l'évolution de la maladie (GMQ, diarrhée,
). Dans cette étude, malgré un effet bénéfique au niveau intestinal, l'incidence d'une supplémentation en vitamine E semble minime pour accroître la résistance des animaux à la colibacillose. |
|
|
|
|
|
Suite
de l'analyse des communications de Pathologie & Hygiène
par P. Coudert 
|
|
|
|
|
|
|
|
|
| |
[1]
Agnoletti F., Favretti M., Deotto S., Passera A., Tisato E., Bano L..,
Mazzolini E., 2004. Report on entheropathogenic Escherichia coli
(EPEC) isolated from enteric ouitbreak in Italian intensive rabbit herds.
Proceedings of the 8th World Rabbit Congress, Puebla (Mexico) Sept.
2004, WRSA ed., 416-421.
[2]
Badiola A., Pérez De Rozas M., Roca M., Carabaño R.,
Garcia J., Rosell J ., 2004. The 16s r-DNA RFLP profile of total DNA
of intestinal bacteria under pathological conditions. Proceedings of
the 8th World Rabbit Congress, Puebla (Mexico) Sept. 2004, WRSA ed.,
427-432.
[3]
Bohez L., Maertens L., Laevens H., Stakenborg T., Peeters J., Vandekerchove
D. , 2004. Use of a 3-/O15 D eae enteropathogenic Escherichia
coli vaccine in a rabbitry with mixed enteropathy problems: spreading
characteristics and protective effect. Proceedings of the 8th World
Rabbit Congress, Puebla (Mexico) Sept. 2004, WRSA ed., 439-444.
[4]
Bohez L., Stakenborg T., Laevens H., Peeters J., Vandekerchove D.,
2004. An attenuated 2+/O132 D tir enteropathogenic Escherichia
coli (EPEC) offers cross protection against a 3-/O15 challenge and
partial protection against an 8+/O103 challenge. Proceedings of the
8th World Rabbit Congress, Puebla (Mexico) Sept. 2004, WRSA ed., 445-450.
[5]
Bohez L., Stakenborg T., Laevens H., Peeters J., Vandekerchove D.,
2004. Different administration methods for the 3-/O15 D eae
EPEC vaccine strain protecting meat rabbits against a 3-/O15 challenge:
preliminary results. Proceedings of the 8th World Rabbit Congress,
Puebla (Mexico) Sept. 2004, WRSA ed., 451-456.
[6]
Boisot P., Duperray J., Guyonvarch A., Richard A., Licois D., Coudert
P. , 2004. Evaluation of the effectiveness of soluble bacitracin (Bacivet
S ®) in drinking water compared to bacitracin in the feed (Albac®),
during an experimental reproduction of epizootic rabbit enteropathy syndrome.
Proceedings of the 8th World Rabbit Congress, Puebla (Mexico) Sept.
2004, WRSA ed., 457-462.
[7]
Boullier S., Nougayrède J-P., Marches O., Tasca C., Milon A.
, 2004. Genetically engineered enteropathogenic Escherichia coli
strain protects rabbits against colibacillosis. Proceedings of
the 8th World Rabbit Congress, Puebla (Mexico) Sept. 2004, WRSA ed.,
463-469.
[8]
Camarda A., Pennelli D., Battista P., Martella V., Greco L., Alloggio
I. , Mazzolini E., 2004. Virulence genes and antimicrobial resistance
patterns of enteropathogenic Escherichia coli from rabbits in
southern Italy . Proceedings of the 8th World Rabbit Congress, Puebla
(Mexico) Sept. 2004, WRSA ed., 470-476.
[9]
Cerrone A., Mariani F., Ciabrelli M., Galiero G., De Carlo E., Fioretti
A., Baiano A., Bartoli M. , 2004. A survey of zoonotic agents in italian
rabbit slaughterhouses. Proceedings of the 8th World Rabbit Congress,
Puebla (Mexico) Sept. 2004, WRSA ed., 490-497.
[10]
Coudert P., Licois D. , 2004. Study of early phenomena during experimental
epizootic rabbit enteropathy: preliminary results. Proceedings of the
8th World Rabbit Congress, Puebla (Mexico) Sept. 2004, WRSA ed., 520-525.
[11]
D'Incau M., Pennelli D., Pacciarini M., Maccabiani G., Lavazza A., Tagliabue
S. , 2004. Characterization of E.coli strains isolated from
rabbits with enteritis in Lombardia and Emilia Romagna (north italy) during
the period 2000-2003. Proceedings of the 8th World Rabbit Congress,
Puebla (Mexico) Sept. 2004, WRSA ed., 526-531.
[12]
Licois D . , 2004. Domestic rabbit enteropathies. Proceedings
of the 8th World Rabbit Congress, Puebla (Mexico) Sept. 2004, WRSA ed.,
385-403 (Rapport invité)
[13]
Pisoni A.M., Piccirillo A., Gallazzi D., Agnoletti F., Grilli G., 2004.
Biotype and susceptibility to antimicrobial agents of rabbit Escherichia
coli. Proceedings of the 8th World Rabbit Congress, Puebla (Mexico)
Sept. 2004, WRSA ed., 608-613.
[14
] Rochambeau H. De, Licois D., Gidenne T., Verdelhan S., Coudert P.,
Elsen J. M. , 2004. Genetic variability of the resistance for three
types of enteropathy in the growing rabbit. Proceedings of the 8th
World Rabbit Congress, Puebla (Mexico) Sept. 2004, WRSA ed., 137-142.
[15]
Tsalie E., Kaldrymidoy E., Kouzi K., Poutahidis Th., Xylouri E., Iliadis
N., Sarris K., Abas Z. , 2004. Ileal mucosal effects of vitamin E
in experimentally infected rabbits with enteropathogenic Escherichia
coli O103. Proceedings of the 8th World Rabbit Congress, Puebla
(Mexico) Sept. 2004, WRSA ed., 639-645.
[16]
Verdelhan S., Bourdillon A., Morel-Saives A., Audoin E. , 2004.
Effect of a limited access to water on mortality of fattening rabbits.
Proceedings of the 8th World Rabbit Congress, Puebla (Mexico) Sept.
2004, WRSA ed., 669-672.
|
|