| |
| I
- MAÎTRISE DE LA REPRODUCTION DE LA LAPINE |
|
1
- Mécanismes physiologiques contrôlant la reproduction
chez la lapine (synthèse C. Boiti - Italie).
Chez la femelle,
la fonction de reproduction englobe différents processus;
du développement folliculaire à l'ovulation, de la
fécondation à l'embryogenèse, de l'implantation
à la mise bas et la lactation. Toutes ces fonctions sont
sous contrôle hormonal. Le point est fait sur les connaissances
acquises ces dernières années chez la lapine au niveau
des mécanismes liés aux aspects nutritionnels, au
stress et au niveau des mécanismes lutéotrophiques
et lutéolytiques.
|
|
|
Mécanismes
liés aux aspects nutritionnels. La leptine intervient
dans la régulation du poids et de la masse graisseuse, elle
contrôle aussi la fonction de reproduction. Cette hormone
est sécrétée principalement par les adipocytes
mais aussi dans de nombreux tissus de l'appareil reproducteur de
la lapine (ovaire, endomètre, placenta et glandes mammaires).
La présence de récepteurs de la leptine au niveau
de l'ovaire, l'oviducte, l'hypothalamus et l'hypophyse antérieure,
suggère que la leptine intervient dans la régulation
aux différents niveaux de l'axe hypothalamo-hypophysaire-ovarien
(HPO). Ainsi, 48 heures de jeûne avant l'insémination
déprime tous les caractères de reproduction, réduit
la fréquence et l'amplitude des pulses du 17 béta-oestradiol,
déprime le pic de LH consécutif à une injection
de GnRH et s'accompagne d'une diminution de la concentration plasmatique
de la leptine. Ce résultat original présenté
au cours de ce congrès (cf partie 3) suggère que la
leptine agit donc comme un signal métabolique qui "allume
ou éteint" l'activité reproductrice.
Avec la leptine, une liste importante de métabolites
tels que le glucose, les acides gras non essentiels, les triglycérides
et des hormones telles que l'insuline et les IGF-I exercent une
effet positif sur l'ovulation, la production de gonadotrophines
et de stéroïdes.
|
 sécrétion
PROLACTINE sécrétion
PROLACTINE

 croissance folliculaire
croissance folliculaire
stéroïdogénèse
RÉCEPTIVITÉ
FERTILITÉ |
Par rapport
aux techniques utilisées sur le terrain pour induire la réceptivité
des lapines au moment d'une insémination artificielle, il
a été montré que 48 heures de séparation
de la mère allaitante et sa portée, avant le moment
prévu pour une insémination s'accompagnent d'une diminution
du niveau de prolactine 24 heures après le début de
la séparation. Cette diminution est suivie d'une augmentation
de la concentration de 17 béta-oestradiol le jour de l'insémination
(48h de séparation) et d'une augmentation du pic de LH déclenché
par l'injection de GnRH. Ce résultat suggère que la
diminution de la sécrétion de prolactine, en relation
avec la suppression temporaire des tétées, a un rôle
clé sur la stimulation de la croissance folliculaire et la
stéroïdogénèse au cours des 48 heures
de séparation et donc sur l'amélioration de la réceptivité
et de la fertilité.
|
| |
Mécanismes
liés au stress. Le stress est susceptible d'activer l'axe
HPO. Le lapin étant très sensible aux situations de
stress, une action permissive ou restrictive peut être observée
sur l'axe HPO en fonction du type et de l'intensité du stress.
Par exemple, des mécanismes liés au stress sont probablement
impliqués quand les lapines sont exposées à des
stimulations environnementales ou d'origine sociale (changement de
cage par exemple) pour améliorer leurs performances de reproduction.
|
48
heures de séparation mère-jeunes
(9e au 11e jour d'allaitement)
- réduction sensibilité au stress
- réduction mortalité nais.-sevr.
- meilleure fertilité de la lapine
|
Parmi les biostimulations
utilisées, il a été montré qu'une courte
séparation entre la mère et les jeunes influence les
systèmes endocriniens, nerveux et immunitaires et en conséquence
le développement post natal. En effet, une courte privation
néonatale maternelle des lapereaux s'accompagne d'une diminution
de la sensibilité au stress et de la mortalité au
sevrage, ainsi que d'une augmentation de la fertilité ultérieure
de la lapine.
|
| |
Mécanismes
lutéotrophiques et lutéolytiques. Des corps jaunes
actifs (activité lutéale) affectent négativement
la fertilité des lapines. En effet, des lapines pseudogestantes
(à la suite d'une
ovulation provoquée en l'absence de la semence d'un mâle
entraînant la présence de corps jaunes sur les ovaires
et une concentration plasmatique de progestérone élevée
mais l'absence d'embryons) ovulent normalement après
une insémination (avec traitement avec GnRH), mais la fécondation
est totalement inhibée. Or des corps jaunes sont parfois
observés sur les ovaires de lapines allaitantes qui n'ont
été ni saillies ni inséminées. Les causes
de ces ovulations primaires ne sont pas connues à ce jour
(ovulation spontanée ?, corps jaunes persistants liés
à une inhibition du mécanisme lutéolytique
? ou problèmes pathologiques ?). Au cours de l'activité
lutéale, plusieurs forces lutéolytiques et lutéotrophiques
s'opposent et agissent sur les corps jaunes. Chez la lapine, le
rôle de l'oxyde-nitrique-synthétase (NOS) dans le fonctionnement
du corps jaune vient d'être mis en évidence. Cet enzyme
serait responsable de l'action lutéotrophique de la prostaglandine
PGE2 ou lutéolytique d'une autre prostaglandine PGF2-alpha
(responsable de la régression des corps jaunes).
|
| |
Malgré
ces avancées, chez la lapine, de nombreux mécanismes
physiologiques sont encore obscurs :
- quels sont les mécanismes intervenant sur l'axe hypothalamo-hypophysaire-ovarien
? Si la leptine participe au contrôle de la fonction de reproduction
chez la lapine, le rôle d'autres hormones du métabolisme
et celui d'autres métabolites doivent être élucidés.
- quels sont les mécanismes liés au stress ?
- quels sont les mécanismes qui contrôlent la durée
de vie des corps jaunes ?
|
| Pour
la souche employée (type Néo Zélandais) les meilleures
performances lors de la première portée sont obtenues
avec des lapines rationnées pendant l'engraissement puis inséminées
à l'âge de 17,5 semaines (portées de 6,4 à
7,8 lapereaux selon les essais) |
2
- Conduite des jeunes lapines pendant leur croissance (synthèse
J. Rommers- Pays Bas)
L'auteur
a testé différentes stratégies d'élevage
des lapines à l'engraissement, susceptibles d'améliorer
leurs performances de reproduction ultérieures ainsi que
leur longévité, et a synthétisé son
travail dans ce rapport invité.
Pendant la phase d'engraissement, la croissance est parfois considérée
comme le facteur majeur qui influence la reproduction ultérieure
des lapines. Dans ces travaux, la croissance est étudiée
en relation avec le niveau alimentaire (ad libitum vs restreint,
niveau de restriction non précisé) et l'âge
à la première insémination (14,5 vs 17,5 semaines).
Les effets des conditions de croissance sont analysés en
fin d'engraissement par le poids et la composition corporelle des
lapines futures reproductrices.
Les lapines restreintes et inséminées à l'âge
de 14,5 semaines sont souvent impubères (seules 50 % ovulent)
et leur poids est faible (3,2 kg). A 14,5 semaines, parmi les lapines
nourries ad libitum, 70 % n'atteignent pas 4 kg [ndlr
: il s'agit de lapines d'une souche de Néo Zélandais].
La compétition entre les besoins de croissance et la production
conduisent donc à des portées plus petites et des
production laitières plus faibles que pour les lapines restreintes
inséminées à 17,5 semaines. En conséquence,
la taille de portée est réduite de 1,4 lapereau. De
plus, les lapines nourries ad libitum inséminées
à 14,5 semaines accroissent leur poids au cours de leur première
gestation et lactation.
A l'âge de 17,5 semaines, plus de 75 % de lapines alimentées
ad libitum pendant l'engraissement pèsent au moins
4 kg. Cependant, les lapines lourdes sont grasses, leur consommation
est plus faible pendant la première gestation et la mortinatalité
augmente. De 60 à 80 % des lapines restreintes inséminées
à 17,5 semaines pèsent 4 kg, le nombre de nés
vivants est plus élevé que chez les lapines nourries
ad libitum. La production laitière est influencée
par la stratégie alimentaire au cours de l'engraissement.
Ainsi les lapines restreintes inséminées à
17,5 semaines produisent plus de lait que celles nourries ad
libitum et inséminées au même âge.
Les lapines restreintes n'ont pas formé de dépôt
de gras excessif à 17,5 semaines et leur consommation alimentaire
est plus élevée que celle des lapines toujours nourries
ad libitum.
|
| A
long terme, aucune différence n'est mise en évidence
entre les différentes stratégies de préparation
des lapines futures reproductrices. |
L'auteur conclut
que, pour obtenir des tailles de portée optimales, les jeunes
lapines doivent peser autour de 4 kg au moment de leur première
insémination. La restriction alimentaire au cours de la phase
d'engraissement permet une meilleure homogénéité
des poids des lapines à la première insémination
et stimule la consommation au moment de leur première gestation.
A la première mise bas, les meilleures performances de reproduction
sont obtenues par les lapines restreintes pendant l'engraissement
et inséminées à 17,5 semaines. Cependant, ces
stratégies d'engraissement n'influencent le poids et la consommation
que pour la parité 1. Des effets à plus long terme
(3 portées) n'ont été mis en évidence
ni sur les performances de reproduction, ni sur le taux d'élimination
des lapines.
|
| |
L'équipe
hongroise de Gyovai et al. a aussi étudié
l'incidence de différentes stratégies de conduite des
lapines futures reproductrices sur leur performances ultérieures
de reproduction. Utilisant un schéma factoriel 3x2x2x2, des
lapereaux nés vivants ont été divisés
en 3 groupes selon leur poids à la naissance (faible 35-45g
, moyen 53-58g, élevé 65-70g) et laissées dans
des portées égalisées à 8. La moitié
des portées a été allaitée par une mère
alors que l'autre moitié des portées a été
nourrie par 2 lapines jusqu'au 17e jour. Une semaine après
le sevrage (21 jours), la moitié des jeunes lapines ont été
nourrie ad libitum, alors que l'autre moitié a reçu
une alimentation restreinte (80-85 % alimentation ad libitum
par limitation du temps d'accès à la mangeoire). Ces
jeunes lapines ont été à nouveau alimentées
à volonté 4 jours avant leur 1ère
insémination. Les jeunes lapines de ces différents groupes
ont été à nouveau divisées en 2 lots et
inséminées soit à 15,5 soit à 18,5 semaines.
L'expérience n'est pas terminée. L'analyse préliminaire
d'un total de 2239 portées issues de 496 femelles conduit aux
conclusions provisoires suivantes : |
| |
- Le poids
à la naissance des jeunes lapines influence leur poids
à tous les stades (3,93 - 3,98 et 4,08 kg en moyenne au
stade adulte). Il est sans effet sur la fertilité, la taille
de portée à la naissance ou au sevrage (7,6 lapereaux
en moyenne à 35 jours), la mortalité naissance-sevrage
ou le poids de portée à 21 jours. Les lapines nées
les plus lègères sèvrent toutefois des portées
de poids plus modeste (6,0 vs 6,3 et 6,2 kg)
- Les lapines
allaitées par 2 mères dans leur jeune âge
sont plus lourdes quand elles sont adultes (4,09 vs 3,92 kg) et
plus prolifiques (7,23 sevrés /MB vs 6,92).
- L'alimentation
ad libitum à partir de 28 jours améliore
le poids à la 1ère insémination mais ensuite
le poids des lapines du groupe restreint est le plus élevé
(conforme aux résultats de Rommers).
- La taille
moyenne des portées à la naissance est plus élevée
chez les lapines inséminées pour la 1ère
fois à 15,5 semaines, ce qui est opposé aux résultats
de Rommers. Mais cet écart a toutefois disparu au sevrage.
Les auteurs
concluent que l'allaitement des futures reproductrices par deux
femelles suivi d'une restriction alimentaire (80-85 % alimentation
ad libitum) jusqu'à la 1ère
insémination a un effet bénéfique sur les performances
ultérieures des lapines.
NDLR
: on peut aussi remarquer, ce qui n'a pas été relevé
l'auteur, que les lapines les plus légères à
la naissance ont donné en moyenne 5,25 portées pendant
l'essai, alors que leurs sœurs nées avec un poids moyen
ou élevé n'en ont donné que 4,34 et 4,10 respectivement.
Au final, chacune des 139 lapines nées légères
ont sevré 37,5 lapereaux pendant l'essai contre 30,5 et 28,9
pour leurs soeurs de poids à la naissance moyen (n=182) ou
élevé (n=175). Il y a matière à réflexion,
... en attendant la fin de l'expérimentation.
|

Lapin "Pannon White" |
Préparation
des futures reproductrices en fin d'engraissement. Milisits
et Lévai (Hongrie) ont comparé les caractéristiques
de reproduction de lapines "Pannon White" dont les réserves
lipidiques ont été estimées (TOBEC) à
l'âge de 10 semaines. Seize pour cent des lapines les plus
grasses (et les plus maigres) ont été accouplées
avec les huit pour cent de mâles ayant les réserves
lipidiques les plus élevées (et les plus faibles).
Les lapines les plus grasses sont plus fertiles mais produisent
des portées plus petites à la naissance. Cependant,
du à une plus faible mortalité avant sevrage, elles
ont des portées plus grandes et plus lourdes à 21
jours que celles ayant des réserves lipidiques plus faibles.
Néanmoins, cette expérience ne permet pas de déterminer
un niveau de réserves lipidiques optimal pour une production
durable et efficace.
|
|
Il semble bien
que le freinage de la croissance par une restriction alimentaire
combiné avec une première insémination tardive
soit favorable à la carrière de reproduction des lapines.
|
Par rapport
à une alimentation ad libitum et une première
insémination des lapines à 16,5 semaines, Bonanno
et al. (Italie) ont montré qu'un régime alimentaire
restreint appliqué à partir de l'âge de 11 semaines
(75 % ad libitum) combiné à un recul de la
première insémination de 3 semaines (19,5 semaines)
conduit à une meilleure composition corporelle des lapines
au moment de l'insémination (poids plus élevé
sans augmentation de la masse lipidique). Cette conduite d'élevage
s'accompagne de l'amélioration de la fertilité des
allaitantes, de la prolificité et plus généralement
de la productivité ultérieure (+ 1,8 kg de lapereaux
produit sur une période de 160 jours) et de la longévité
(taux de fonte du cheptel de 48,5% après 160 jours de production
contre 54,8% pour le lot témoin ad libitum avec 1ère
insémination à 16,5 semaines). Il faut cependant ajouter
que les lapines restreintes ont reçu un flushing alimentaire
10 jours avant la première l'insémination et sur l'ensemble
des lapines, un allaitement contrôlé était pratiqué
les 2 jours précédant les inséminations ultérieures.
|
| Les
lapines reproductrices sont peu sensibles à un apport excessif
de sodium |
Par
ailleurs, l'utilisation prolongée (de la 1ère
saillie, jusqu'au sevrage de la 3ème
portée) d'un aliment avec un fort apport de sodium (35 meq/100g
vs 27meq/100g de Na+K-Cl obtenus par addition de bicarbonate de
sodium à une ration de base contenant déjà
1,5% de NaCl), n'affecte ni les profils métaboliques ou minéraux,
ni la concentration plasmatique d'œstradiol, mais a tendance
à augmenter la sécrétion de cortisol (P<0,10)
chez les lapines multipares et allaitantes. Les hormones thyroïdiennes
tendent à augmenter mais les écarts ne sont pas significatifs
(Chiericato et al.- Italie). Les auteurs mentionnent que
leurs résultats antérieurs avaient montré l'absence
d'effet de tels apports de sodium sur les performances de reproduction.
|
| |
Ces travaux
montrent à minima, qu'il existe avant et après la
période globale d'engraissement (du sevrage à la 1ère
IA) des stratégies susceptibles d'améliorer et sans
doute d'homogénéiser les performances de reproduction
ultérieures des reproductrices. Malgré quelques résultats
parfois contradictoires (génotypes et conditions expérimentales
divers), il semble qu'un rationnement des animaux pendant tout ou
partie de la phase d'engraissement, combiné à une
insémination pratiquée au plus tôt à
17,5 semaines, permet d'améliorer la composition corporelle
à la 1ère insémination et d'améliorer
les performances de reproduction des reproductrices.
|
| |
3
- Biostimulations et induction de la réceptivité sexuelle
des lapines
La recherche
d'alternatives à l'utilisation éventuelle d'hormones
pour induire la réceptivité sexuelle des lapines a
pour objectif d'améliorer et d'homogénéiser
les performances de reproduction dans un contexte "d'agriculture
durable" qui prend en compte la santé des animaux, leur
bien être ainsi que la santé humaine. Le programme
européen Cost 848 a permis de favoriser les échanges
entre les différentes équipes européennes engagées
sur ce thème, en particulier dans le cadre de l'International
Rabbit Reproduction Group. Trente pour cent des communications
de la session Reproduction concernent ce thème. Sous le terme
de "biostimulation", nous comprenons toute stimulation
environnementale appliquée les jours précédant
l'insémination.
|
| Une
restriction alimentaire d'une ou deux journées avec reprise
de l'alimentation 2 heures avant une insémination n'a que des
conséquences négatives sur la fonction de reproduction |
Restriction
alimentaire. Brecchia et al. ont étudié
les effets d'un jeûne de 24 ou 48 heures avant l'insémination
sur l'axe ovarien et les performances de reproduction des lapines.
Le challenge nutritionnel (biostimulation) consistait à alimenter
à nouveau les lapines 2 heures avant l'insémination.
Par rapport à un groupe de lapines nourries ad libitum,
le jeûne appliqué durant 1 à 2 jours avant l'insémination
déprime la réceptivité, la fertilité
et le nombre de nés vivants. Les conséquences du jeûne
sur l'axe hypothalamo-hypophysaire-ovarien sont : la diminution
de l'expression des récepteurs des œstrogènes
au niveau hypothalamo-hypophysaire, de la fréquence et de
l'amplitude des pulses des œstrogènes, de l'amplitude
du pic LH 30 à 60 minutes après l'injection de GnRH
et de la concentration plasmatique de la leptine en fin de jeûne.
Le flushing alimentaire pratiqué 2 heures avant l'insémination
a donc été insuffisant. Cependant, ce travail permet
une meilleure compréhension des relations entre nutrition
et reproduction au travers de l'axe HPO.
Gomez
et al. (Mexique) ont comparé les effets de différentes
biostimulations ou restrictions, sur la réceptivité
de lapines nullipares Californiennes ou Néo-zélandaises:
flushing, jeûne de 24h avant saillie, changement de cage.
Quel que soit le génotype, 24 heures de jeûne avant
la première saillie, dépriment la réceptivité
des lapines le jour de la présentation. Par contre, l'effet
de la stimulation (ou restriction) peut dépendre du génotype
des lapines. Ainsi par exemple, un changement de cage 24h avant
la saillie, n'améliore la réceptivité que des
lapines californiennes, et l'effet du flushing (200 g d'aliment
par jour, les 5 jours précédant la saillie, au lieu
de 100 g distribués jusque là) est favorable chez
les lapines Californiennes (78% vs 32% de lapines réceptives)
et défavorable chez les lapines Néo Zélandaises
(35% de réceptives vs 62%)
|
| Un
allaitement effectué sous contrôle entre 8h45 et 9h00
du 9e au 13e jour après une mise bas permet d'obtenir un meilleur
taux de réussite des inséminations (pratiquées
le 11e jour 15 mn après la fin de l'allaitement), que celui
d'inséminations faites au même moment chez des lapines
libres d'allaiter leur portée. Les tailles de portées
peuvent aussi être améliorées. |
Séparation
mère-jeunes. Eiben et al. (Hongrie) ont étudié
la productivité des lapines soumises à un allaitement
contrôlé de 2 à 5 jours autour de l'insémination.
Par rapport à un lot témoin (allaitement libre), 2
jours d'allaitement contrôlé avant l'insémination
poursuivis 3 jours après l'insémination permettent
d'améliorer significativement la productivité des
lapines, mais il faut mentionner que le lot témoin avait
une fertilité particulièrement basse (33% d'IA fécondes
contre 60% dans le lot avec allaitement contrôlé).
Par ailleurs, Matics et al. (Hongrie-France) ont mesuré,
par rapport à un lot témoin conduit en allaitement
libre, l'effet d'un allaitement contrôlé pratiqué
2 ou 3 jours avant l'insémination. Ils observent une meilleure
réceptivité (d'après la couleur de la vulve)
mais sans gain sur la fertilité (78 à 80% de lapines
fécondées après IA pour les 3 lots). Cependant,
contrairement aux résultats d'Eiben et al., le nombre
de nés vivants est amélioré quand les lapines
sont soumises à 3 jours d'allaitement contrôlé
avant l'insémination. Cette stimulation se traduit par un
gain de productivité de 19 % de nés vivants par insémination
réalisée.
Sur un plan physiologique, Rebollar et al. (Espagne) ont
montré sur des lapines allaitantes (4 jours post partum),
que 48 heures de séparation de leur portée, s'accompagne
d'une augmentation de la concentration plasmatique d'œstradiol
(par rapport aux lapines non stimulées) mais pas d'une diminution
de prolactine comme l'avaient montré Ubilla et al.
sur des lapines allaitantes au stade 11 jours post partum.
|
Le fractionnement
de la période d'éclairement de 16 h en 2 fois 8h
ne présente aucun avantage
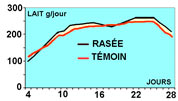
Figure 1 : Production laitière de lapines rasées
ou non 2 jours avant l'IA (10 j post partum)
|
Programmes
lumineux. Au cours de ce congrès, 2 communications avaient
pour objectif d'étudier des programmes lumineux de base sur
lesquels des recherches ultérieures pourraient s'appuyer,
pour étudier l'effet de stimulations lumineuses destinées
à induire la réceptivité sexuelle des lapines
au moment de l'insémination. Szendrö et al. (Hongrie)
ont comparé les performances de reproduction de lapines soumises
dès 11 semaines à 16 heures d'éclairement quotidien
distribué de façon continue (16L:8D) ou discontinue
(8L:4D:8L:4D). Sous 16 heures d'éclairement fractionné,
le poids des lapines est plus élevé à la mise
bas. Cependant, ni la fertilité, ni la taille de portée,
ni la viabilité des jeunes, ni le poids de portée
ou le poids individuel à 21 jours ne sont influencés
par le type de distribution de la lumière dans des conditions
d'utilisation de l'allaitement contrôlé.
Les systèmes d'élevage ayant fortement évolué
ces quinze dernières années, Theau-Clément
et al. (France) ont voulu vérifier la pertinence du
choix des programmes lumineux utilisés dans les élevages
et étudier s'il existe une variabilité génétique
de la sensibilité au photopériodisme. Les performances
de reproduction de lapines issues de 2 types génétiques
(génotype 67 type hybride commercial et génotype 57croisement
expérimental incluant des lapins Rex) ont été
étudiées. Elles étaient soumises à 3
programmes lumineux : 8 heures d'éclairement quotidien continu
(8L:16D), ou 16 heures d'éclairement continu (16L:8D) ou
discontinu (8L:4D:8L:4D). Un total de 1548 inséminations
ont été réalisées au cours de 8 séries
dans des conditions d'allaitement libre. Le programme lumineux n'influence
pas la durée de production des lapines. Sous seize heures
d'éclairement continu, les lapines 67 sont plus réceptives.
Les tailles de portée à la naissance et au sevrage
sont plus élevées sous 16 heures d'éclairement
(continu ou discontinu) quel que soit le type génétique,
cependant, les lapereaux issus de mères 67 sont plus lourds.
A l'opposé, les lapines sont moins fertiles sous 16 heures
de lumière (74-76% vs 83% avec 8L:16D). En conséquence,
les programmes lumineux influencent peu la productivité globale,
ils n'interagissent pas avec le génotype des femelles sur
la fertilité et la prolificité. Cependant, sur les
femelles de production de chair (67), il est confirmé que
16 heures d'éclairement continu peuvent être recommandés
dans la mesure où ce programme s'accompagne d'une augmentation
de la réceptivité des lapines et de la croissance
des jeunes sous la mère. Ces 2 dernières études
montrent que, quelles que soient les conditions d'allaitement, 16
heures d'éclairement discontinu n'améliorent pas,
par rapport à 16 heures d'éclairement continu, les
performances de reproduction des lapines.
Rasage des lapines en saison chaude. Dans l'objectif de diminuer
le stress lié aux fortes chaleurs et d'améliorer les
performances de reproduction l'été, Szendrö et
al. (Hongrie) ont mesuré l'effet du rasage de la partie
dorsale et des flancs des reproductrices, 2 jours avant l'insémination.
Cette pratique améliore la production laitière et
le poids des portées à 35 jours. Cependant, elle n'influence
ni la fertilité, ni la prolificité. Si cette stimulation
permet d'améliorer la croissance des lapereaux, elle est
insuffisante pour améliorer les performances de reproduction
des lapines.
|
| |
Une
restriction alimentaire totale (jeûne 24-48 h avant IA) s'accompagne
d'une altération systématique des performances de reproduction
des lapines. A l'inverse, on ne connaît pas aujourd'hui de technique
de "flushing alimentaire" pratiqué quelques jours
avant l'insémination, susceptible d'améliorer régulièrement
les performances des lapines inséminées. Une séparation
mère-jeunes de 36 heures ou un allaitement contrôlé
pratiqué 3 jours avant l'insémination, éventuellement
poursuivi 2 jours après, permettent d'améliorer la productivité
des lapines. Dans les élevages de production de chair, il est
confirmé que 16 heures d'éclairement continu peuvent
être recommandés. Il convient maintenant d'étudier
des techniques de stimulation lumineuses susceptibles d'induire la
réceptivité sexuelle des lapines et d'améliorer
leur productivité ainsi que la viabilité et la croissance
des jeunes. |
| |
4
- La capacité utérine et diverses études sur
la reproduction de la femelle
Sur
des lapines issues d'une sélection divergente sur la capacité
utérine, Argente et al. (Espagne) ont mesuré
sur des lapines hémiovariectomisées (suppression d'un
ovaire) ou non, l'espace disponible pour les fœtus et leur
développement à 18 jours de gestation. Le taux d'ovulation
de l'ovaire restant est double chez les lapines hémiovariectomisées
(12,4 vs 6,5 corps jaunes pour 1 seul ovaire de lapine intacte).
Les résultats suggèrent que chaque embryon a besoin
d'un espace utérin minimal pour s'implanter, survivre et
se développer. La position fœtale dans l'utérus
n'affecte pas la survie des embryons. Cependant, les sites d'implantations
alimentés par moins de 3 vaisseaux ont des placentas et des
fœtus plus légers (respectivement 1,31 vs 1,41g
et 2,03 vs 2,12 g) que ceux alimentés par plus de
3 vaisseaux. De même, les fœtus ayant des placentas recevant
moins de 3 vaisseaux ont une probabilité de mortalité
supérieure (35 % vs 4 %). Ces observations conduisent
à l'identification de caractères qui pourraient être
sélectionnés pour améliorer et homogénéiser
la productivité des lapines.
Gutierrez
et Zamora (Mexique) ont étudié la viabilité
embryonnaire à différents stades de gestation en comparant
les observations obtenues par échographie avec des mesures
in vivo après abattage des lapines. L'échographie
permet de détecter la gestation dès 7 jours mais ne
permet pas de mesurer toutes les vésicules embryonnaires
et donc de dénombrer les embryons.
|
| Des
embryons peuvent parfois se développer en dehors de l'utérus,
provoquant une gestation extra-utérine. |
Dans la session pathologie, l'étude de Corpa et al.
(Espagne) doit être remarquée. Il y a gestation extra-utérine
ou abdominale quand l'implantation et le développement d'un
ou plusieurs oeufs segmentés a lieu dans la cavité abdominale.
Ce phénomène est en principe rare [0,7%
des observations d'autopsie selon Rosell (2000)]. Corpa et
al. distinguent la gestation abdominale primaire, si aucune rupture
utérine n'est observée, supposant que les embryons ont
été expulsés dans la cavité abdominale,
de la gestation abdominale secondaire quand une rupture utérine
est observée. Sur 550 observations de lapines réformées,
issues de 2 élevages de race Néo Zélandaise,
ces auteurs ont identifié 28 gestations abdominales (7 primaires
et 22 secondaires, soit au total 5,1% des lapines observées).
Les auteurs concluent que les causes de cette pathologie pourraient
être étudiées en liaison avec les nouveaux systèmes
d'élevage. En effet, le taux de gestation extra-utérine
est plus élevé dans l'élevage conduit en insémination
que dans celui conduit en saillie naturelle (7,8 % vs 1,6 %).
[ndlr : avant de conclure,
il conviendrait d'abord de vérifier la fréquence de
ce phénomène sur un beaucoup plus grand nombre d'élevages,
et pour les lapines en IA vérifier le rôle de l'inséminateur] |
| |
|
| |
|
|
|